Теория многокомпонентного цветного зрения


А. Фоторецептор-колбочка с пигментами. Свет проходит сквозь хрусталик и фиксируется (фильтруется) соответствующим пигментом в конусной мембране колбочки, расположенной на её «дне». Как видим имеются четыре сечения мембраны с пигментами четырёх цветов. (У человека три сечения цветов пигмента RGB — трихроматизм). Колбочка А с четырьмя вариантами работы при её освещении.
Б.1-Б.4 У птиц обычно четыре варианта работы колбочки (они «тетрахроматы»), что позволяют им синтезировать многоцветное изображение, и различать цвета лучше, чем человек. Обработка полученных разностных сигналов, происходящая в нейронной сети сетчатки (десятки типов клеток) обеспечивает возможность ясного различения тысяч цветов и оттенков. Важно, что схема цветного зрения человека может быть показана в виде колбочки A, способная принимать любой луч основного цвета RGB в трёх соответственных сечениях внешней мембраны S,M,L — трихроматизма. В зависимости от расположения колбочки её внешняя мембрана может принять красные, зелёные лучи (M,L) в зоне фовея, лучи S,M,L в периферийной части сетчатки, а в наиболее удалённых поясах сетчати — (S — синие лучи спектра). Работа мембран фоторецепторов сетчатки глаза как волновод физика Медейроса, Специфика работы мембраны колбочек сетчатки глаза.[2][3]



.jpg)
Многокомпонентная теория цветного зрения — теория, объясняющая цветовое зрение человека на базе (трихроматизма), которая основана на принципах биофизики, биохимии, физики, что существуют три «основных» видов цвета RGB, которые при смешивании дают возможность получить любой оттенок цветов, например, как это делает художник.
Среди животных зрительная система работает на принципах четырёхроматизма, пяти и более. (См. ниже.)
На базе физики цвета (RGB) и взаимодействия с ним живой среды обитания — биологической, а также на основе колориметрических, гистологических, генетических исследований,[3] на базе достижений в этой области более, чем за 30‒35 лет, на основе биофизики, биохимии, пересмотра основ прежнего сложившегося процесса зрения с разных точек зрения — имеем:
- С точки зрения биологической, в области цветного зрения начиная с 1966 по 2009 годы (Труды доктора Р.Е.Марка и его лаборатории) с основными экспериментальными данными исследований живой клетки, на срезах сетчатки установлена работа колбочек и (S,M,L) в блоке RGB и палочек. В условиях дневного освещения (цветного зрения) — колбочек и работа палочек, в период сумеречного и ночного освещения (не цветного) изолировано от колбочек. Работа фоторецепторов связана с полученными данными работы видоизменяющихся разновидностей фотопигментов на базе белков опсинов:
- Кон-опсины(конопсин) — у колбочек,
- Род-опсин(родопсин) — у палочек.[1]
- С именем Тимоти Голдсмит связано фундаментальное исследования вопросов цветного зрения у птиц[6], которые подтвердили не доказанные до него многокомпонентные теории цветного зрения. Он провёл на гистологическом уровне с применением современного оптического устройства — флуоресцентного микроскопа исследования живых клеток (разрешение 1‒10нм) с получением цветных стереоизображений.
- *С точки зрения чисто физической на базе труда (2011 года) учёного физика[4] Джеральда К. Хата, который на базе физики взаимодействия света с внешними долями мембран фоторецепторов сетчатки глаза, где в основном осуществляется первичное взаимодействие со светом на наноуровне («nanostructural») (рецепторное) и доктора физика Джона Медейроса,[2] который рассмотрел работу внешних долей мембраны колбочек и палочек с точки зрения аналогичной работе волноводов конической и цилиндрической формы в среде прозрачного тела глаза (жидкая среда). В конечном итоге, физики Дж. К. Хат и Джон Медейрос пришли к общепринятому принципу трихроматизма. (См. Пересмотр традиционных взглядов на зрительный процесс физика К. Хата, Работа внешних мембран колбочек и палочек сетчатки глаза как волновод).
- *Данные доктора, физика Джона Медейроса, рассмотрев внешние доли мембран колбочек и палочек, он их рассматривает как биологические волноводы, которые воспринимают световые лучи с фиксацией их в сечениях волноводов, равных размерам поперечного сечения фронта световой волны в обратном порядке прохождения их (внешних долей мембран колбочек) в жидкой среде, нежели в обычных оптических волноводах в воздухе. (Связано с влиянием показателей преломления сред).
- Фундаментальные исследования академика РАН биохимика М. А. Островского[5] в области работы фотопигмента родопсина Биохимия зрения и свободно-радикальное окисление клеток сетчатки глаза (Участие палочек в условиях сумеречного и ночного освещения);
- *Исследования РАН Е. О. Загальская, В. П. Гнюбкина [6] (См. Ретиномоторная реакция фоторецепторов сетчатки глаза);
Сущность теории[править | править код]
В глазу человека содержатся два типа светочувствительных клеток (экстерорецепторов): высокочувствительные палочки и менее чувствительные колбочки. Палочки функционируют в условиях относительно низкой освещённости и отвечают за действие механизма ночного зрения, однако при этом они обеспечивают только нейтральное в цветовом отношении восприятие действительности, ограниченное участием белого, серого и чёрного цветов. Колбочки работают при более высоких уровнях освещённости, чем палочки. Они ответственны за механизм дневного зрения, отличительной особенностью которого является способность обеспечения цветового зрения.
У приматов (в том числе и человека) мутация вызвала появление цветовых рецепторов. Это было вызвано расширением экологической ниши млекопитающих, переходом части видов к дневному образу жизни, в том числе на деревьях. Мутация была вызвана появлением изменённой копии гена, отвечающего за восприятие средней, зелёночувствительной области спектра. Она обеспечила лучшее распознавание объектов «дневного мира» — плодов, цветов, листьев.
У птиц при цветном зрении работабт 4 колбочки, откуда и название статьи носит характер Многокомпонентная теория....


В сетчатке глаза человека есть три вида колбочек, максимумы чувствительности которых приходятся на красный, зелёный и синий участки спектра.[1][7] Ещё в 1970-х годах было показано, что распределение типов колбочек в сетчатке неравномерно: «синие» колбочки находятся ближе к периферии, в то время как «красные» и «зеленые» распределены случайным образом,[2] что было подтверждено более детальными исследованиями в начале XXI века.[3] Соответствие типов колбочек трём «основным» цветам обеспечивает распознавание тысяч цветов и оттенков. Кривые спектральной чувствительности трёх видов колбочек частично перекрываются, что способствует явлению метамерии. Очень сильный свет возбуждает все 3 типа рецепторов, и потому воспринимается, как излучение слепяще-белого цвета (эффект метамерии).
Равномерное раздражение всех трёх колбочек, соответствующее средневзвешенному дневному свету, также вызывает ощущение белого цвета (см. Психология восприятия цвета).
Свет с разной длиной волны по-разному стимулирует разные типы колбочек. Например, желто-зелёный свет в равной степени стимулирует колбочки L и M-типов, но слабее стимулирует колбочки S-типа. Красный свет стимулирует колбочки L-типа намного сильнее, чем колбочки M-типа, а S-типа не стимулирует почти совсем; зелено-голубой свет стимулирует рецепторы M-типа сильнее, чем L-типа, а рецепторы S-типа — ещё немного сильнее; свет с этой длиной волны наиболее сильно стимулирует также палочки. Фиолетовый свет стимулирует почти исключительно колбочки S-типа. Мозг воспринимает комбинированную информацию от разных рецепторов, что обеспечивает различное восприятие света с разной длиной волны.
Зрительные пигменты[править | править код]
Установлено [9][10], что в состав зрительных пигментов входят опсины (версия Миг). Различные опсины различаются аминокислотами в составе молекул, и поглощают свет в несколько различном диапазоне длин волн, как ретиналь-связанные молекулы.
Впервые существование колбочкового пигмента (косвенным образом) было обнаружено Джорджем Уолдом, который и дал ему название йодопсин.[11]. В 1967 г. за эти работы ему была присуждена Нобелевскую премию по физиологии и медицине.
Различные опсины различаются аминокислотами в составе молекул, и поглощают свет в несколько различном диапазоне длин волн, как ретиналь-связанные молекулы.
У человека идентифицировано три вида фотопигментов колбочек йодопсина (фотопсина):
- 560 нм: красно-чувствительный L-иодопсин с L-фотопсином. Дефекты гена OPN1LW являются причиной протанопии (красный цвет).
- 530 нм: чувствительный к зеленому M-иодопсин с M-фотопсином. Дефекты гена OPN1MW являются причиной дейтеранопии (зелёный цвет).
- 420 нм: сине-чувствительный S-йодопсин с S-фотопсином. Дефекты гена OPN1SW являются причиной тританопии (синий, УФ цвет).
| Тип колбочки | Название | Диапазон | Пиковая длина волны[12][13] |
|---|---|---|---|
| S (OPN1SW — ген для нормального цветного зрения (S)) — «тритан», «цианолаб» | β | 400—500 нм | 420‒440 нм (УФ, синий) |
| M (OPN1MW — ген для нормального цветного зрения (M)) — «деутан», «хлоролаб» | γ | 450—630 нм | 534‒545 нм (зелёный) |
| L (OPN1LW — ген для нормального цветного зрения (L)) — «протан», «эритролаб» | ρ | 500—700 нм | 564‒580 нм (красный) |
За цветовое зрение человека и обезьян отвечают гены, кодирующие светочувствительные белки опсины. По мнению сторонников трёхкомпонентной теории, наличие трёх разных белков, реагирующих на разные длины волн, является достаточным для цветового восприятия. У большинства млекопитающих таких генов только два, поэтому они имеют двухцветное зрение. В том случае, если у человека два белка, кодируемые разными генами, оказываются слишком схожи или один из белков не синтезируется, развивается дальтонизм.
Чувствительный к красному свету опсин кодируется у человека геном OPN1LW[6].
Другие опсины человека кодируют гены OPN1MW, OPN1MW2 и OPN1SW, первые два из них кодируют белки, чувствительные к свету со средними длинами волны, а третий отвечает за опсин, чувствительный к коротковолновой части спектра.
Необходимость трех типов опсинов для цветового зрения недавно была доказана в опытах на беличьей обезьяне (саймири), самцов которых удалось излечить от врожденного дальтонизма путем введения в их сетчатку гена человеческого опсина OPN1LW[7]. Эта работа (вместе с аналогичными опытами на мышах) показала, что зрелый мозг способен приспособиться к новым сенсорным возможностям глаза.
Ген OPN1LW, который кодирует пигмент, отвечающий за восприятие красного цвета, высоко полиморфен (в недавней работе Виррелли и Тишкова было найдено 85 аллелей в выборке из 256 человек[8]), и около 10 % женщин[9], имеющих два разных аллеля этого гена, фактически имеют дополнительный тип цветовых рецепторов и некоторую степень четырёхкомпонентного цветового зрения.[10] Вариации гена OPN1MW, который кодирует «желто-зеленый» пигмент, встречаются редко и не влияют на спектральную чувствительность рецепторов.
Ген OPN1LW и гены, отвечающие за восприятие света со средней длиной волны, расположены в Х-хромосоме тандемно, и между ними часто происходит негомологичная рекомбинация или генная конверсия. При этом может происходить слияние генов или увеличение числа их копий в хромосоме. Дефекты гена OPN1LW — причина частичной цветовой слепоты, протанопии[6].
Введение[править | править код]
С учётом этих всех вышеперечисленных современных взглядов известных учёных в области биофизики, биохимии, физики пересмотрены основы прежнего сложившегося процесса цветного зрения с разных точек зрения (см. выше) в настоящее время доказана и принята точка зрения, что цветное зрение — это работа экстерорецепторов сетчатки глаза колбочек (без палочек) в связи со всеми остальными клетками сетчатки включая зрительные отделы коры головного мозга.
Фотопигмент с общим названием Йодопсин (версия Миг) содержится в колбочках и участвует в механизме цветового зрения. Согласно многокомпонентной теории цветного зрения в настоящее время принято, что существуют три вида этого пигмента и сетчатка глаза соответственно содержит самостоятельные колбочки RGB, которые в зависимости от попадания на них сфокусированных световых лучей предметных точек с разными длинами волн (S,M,L) трансдукцируют биосигналы в мозг оппонентно отобранные основные лучи видимого спектра света предметных точек: голубого, зелёного и красного цвета RGB. Однако, пока известно, что йодопсин (версия Миг) находится во всех колбочках глаза. Он включает в себя фотопигменты хлоролаб, который поглощает лучи жёлто-зелёной части спектра) и эритролаб, который поглощает лучи жёлто-красной части спектра. На сегодня пока не выделены все типы фотопигментов. Обнаружить присутствие третьего только теоретически предсказанного синечувствительного фотопигмента цианолаб (версия Миг), который (по трёхкомпонентной теории цветового зрения) содержится в колбочке «в третьем случае приёма синих лучей», пока не удалось (ни у дихроматов, ни у трихроматов), что объясняется жёлтой пигментацией в районе центральной ямки, которая по мнению исследователей затрудняет идентифицировать фотопигмент при измерении, отборе, фототранслукции сигнала при воздействии на мембрану колбочек-S лучей фиолетово-синего спектра, которые обладают высокой частотой колебаний элетромагнитных волн и малой временной задержкой в период выделения биосигнала сфокусированной предметной точки в фокальной поверхности сетчатки. (Период в фемтосекундах) (См. Идентификация разновидностей йодопсина — цианолаба).
Тем не менее, получены снимки синей колбочки-S в плане на сетчатке глаза на базе фундаментальных исследования доктора Р.Е.Марка в его лаборатории Труды доктора Р.Е.Марка и его лаборатории [8][9][10], где уже даже без применения красителей легко получили изображение сетчатки с синей колбочкой-S (См. ниже рис. S). Это значит, что пигмент цианолаб в колбочке-s работает. См. ниже — Выводы, где дан более глубокий анализ несостоятельности утверждений сторонниками и автором нелинейной теории цветного зрения по вопросу синей колбочки, и пигментов.
Уже можно утверждать, что в настоящее время не стоит вопрос о принципах теории многокомпонентного цветного зрения в целом. Известные лаборатории в области зрительной системы занимаются в настоящее время вопросами работы всех нервных клеток глаза и мозга, участвуют в создании, проведения биосигнала предметной точки оптического изображения на рецепторном (сетчатка) и нейронном уровне в мозгу. То есть ведётся работа по разработке атласов прохождения сигналов с целью возможности диагностирования, лечения заболеваний зрительной системы. (См. Труды доктора Р.Е.Марка и его лаборатории).
Таким образом имеем :
- Опубликованный аналитический обзор Роберта Марка[11] — 2009 г. (см. также Труды доктора Р.Е.Марка и его лаборатории;
- Труды ученого физика Джеральда К. Хата[12] Пересмотр традиционных взглядов на зрительный процесс физика К. Хата;
- Труды физика доктора Джона Медейроса Работа внешних мембран колбочек и палочек сетчатки глаза как волновод[13];
- фундаментальные исследования академика РАН биохимика М. А. Островского[14] в области работы фотопигмента родопсина Биохимия зрения и свободно-радикальное окисление клеток сетчатки глаза;
- Исследования РАН[15] Ретиномоторная реакция фоторецепторов сетчатки глаза);
Где подтверждено, доказано, что в основе цветного зрения лежит принцип трихроматизма, при дневном освещении работают колбочки, при сумеречном и ночном — палочки. В основе зрительной системы лежат ощущения, связанные с рецепторным восприятием живыми организмами предметных точек под воздействием светового излучения, прямыми лучами источника света, или отражёнными лучами света с последующей дифференциацией ощущений в зависимости от длины волны света. Нервная система получает, сравнивает преобразованные цветовые сигналы — ответы, которые формируются вначале экстерорецепторах (фоторецепорах) — колбочках и палочках в фокальной поверхности сетчатки глаза. При этом участвуют независимые фоторецепторы ipRGC. На рецепторном уровне происходит выделение основных сигналов RGB (S,M,L — «синих», «зелёных», «красных»). Фоторецепторы ipRGC связаны с ними (рецепторный уровень) и участвуют в последующей трансдукции этих биосигналов (бесцветных) в головной мозг. В мозгу происходит окончательное формирование (нейронный уровень) цветного, стерео оптического изображения (в зрительных отделах головного мозга) — появления ощущения цвета.
Что касается других вопросов, то это:
- Создание и совершенствования теорий цветного зрения, которые в общем изложены в статье Теории цветного зрения (версия Миг) и частично — в Истории оптики.
- То, что мы ощущаем как цвет, представляет собой комбинированное воздействие:
- 1) Спектрального распределения светового потока из дающего энергию источника света;
- 2) Физических и/или химических свойств всех материалов, пропускающих или отражающих световой поток (по меньшей мере часть светового потока, переориентированную в сторону глаза);
- 3) Физиологической реакции глаза на световой поток, включающей в себя нервные импульсы, передаваемые в ту часть коры головного мозга, которая отвечает за зрение;
- 4) Переработки нашим мозгом этих сигналов в сочетании с сигналами из соседних областей поля зрения, нашими воспоминаниями о сходных ситуациях, имевших место в прошлом опыте.[14]
- Хотя это достаточно общее высказывание в целом верно, но для того, чтобы «теория цвета» стала продуктивной, требуется дальше развивать и углублять, связывать с фактами каждую модель уровней цветовосприятия.
Требования к теории восприятия света и цвета[править | править код]
Факты и их объяснение[править | править код]
К главным вопросам, раскрывающих суть теории трёхкомпонентного цветного зрения, относятся:
- Экспериментальные гистологические научные данные, проводимые при микроскопии на базе современных Флюоресцентных микроскопов с использованием живых клеток (сечений сетчатки глаза) с разрешающей способностью 1‒10нм, в цветовом трёхмерном пространстве на атомно-молекулярном уровне. (См. Цветное зрение у птиц (версия Миг));
- Колориметрия (наука о цвете);
- Цветоведение;
- Оптика;
- Физиология и анатомия человека;
- Биохимия восприятия света;
- Генетика;
- Психология восприятия цвета.
Все указанные факторы должны быть непротиворечиво объединены с помощью наиболее понятного понятийного и математического аппарата.
Среди фактов, которые должны быть учтены:
- Данные последних исследований строения сетчатки при микроскопии живых клеток (рентгеноскопии, флюоресцентной наноскопии);
- Учёт спектра чисто земных изображений, который может быть в виде двух основных компонентов:
- Цветной контраст между короткими и средними длинами длин волн спектра;
- Цветной контраст между средними и длинными частями длины волны спектра.
- Должна быть использована информация о приматах и человеке, и чтобы извлечь и использовать эту информацию, требуется:
- Определение в видимой части соответствующей области длины волны;
- Учёт морфологии (схемы) сетчатки глаза, которая должна приспосабливаться, воспринять спектральную информацию и быть эффективно переданной в мозг.
Животные, приматы, зрительные системы которых отвечают этим требованиям, наделены способностью цветного зрения.[15][16]
- Должно быть учтено и выдвинуто на первый план недавнее продвижение понимания трех аспектов видения цвета trichromaticу у приматов (2002г):
- 1) поведенческое значение trichromacy,
- 2) развитие и генетика фотопигментов,
- 3) относящиеся к сетчатке глаза кругообороты, которые создают красно-зеленые и синие-желтые цветные каналы в зрительном нерве.[17]
Общие требования[править | править код]
Теория цветного зрения многокомпонентная как и любая теория может считаться полезной только при выполнении следующих условий:
- теория должна строиться на базе объективных, подтверждённых экспериментальных данных,
- на основе использования информации современной микроскопии и гистологического анализа (например, флюоресцентной микроскопии живых тканей) на атомно-молекулярном уровне, в цветовом 3D пространстве,
- теория должна основываться на физических законах,
- научные способы (включая Научное моделирование) должны содержать совокупность основных способов и методов решения задач в рамках получения новых знаний,
- умозаключения и выводы должны получаться с помощью правил и принципов рассуждений на основе эмпирических (наблюдаемых и измеряемых) данных об объекте.[18]
- учёт работы всех фоторецепторов сетчатки и фотопигментов, их нейронные связи между собой и мозгом,
- объяснение основных и дополнительных цветов, понимание их влияния на контрастность, спектральную чувствительности человеческого зрительного восприятия яркости света, изображения,
- законов смешивания цветов (например, аддитивный синтез цвета, субтрактивный синтез цвета),
- основываться на биофизических и биохимические законах,
- описываться математическими зависимостями в трёхмерном 3D пространстве.
Феноменология цветовосприятия[править | править код]

Феноменология цветовосприятия (греч., от phainomenon — феномен, и logos — слово, или наука о явлениях в природе) — когда различные феномены цветного зрения показывают, что зрительное восприятие света, цвета зависит не только от вида воздействующих источников света и цвета и работы фоторецепторов, но также от способа переработки световых сигналов в зрительной системе при зрительной фототрансдукции фоторецепторов сетчатки глаза.
Различные участки видимого спектра света кажутся нам по-разному окрашенными, при этом отмечается непрерывное изменение ощущений восприятия цвета при переходе от фиолетового и синего через зелёный и жёлтый цвета к красному. Одновременно человек может воспринимать цвета, которые не спектральные (отсутствующие в спектре), например, пурпурный цвет, получаемый при смешении красного и синего цветов. Различные физические способы цветового воздействия зрительной системы могут приводить к идентичному восприятию цвета. Например, монохроматический жёлтый цвет не отличается от соответствующей смеси монохроматических зелёного и красного цветов. Или при воздействии на зрительную систему нужным набором цветов RGB,S,M,L, («белого» цвета, например, солнечного) на диске, то при его вращении мы увидим диск белого цвета, и выполнив тоже самое, но c набором XYZ — мы увидим чёрный цвет (точнее тёмно-коричневый) (см. рис.3).
Феноменологию цветовосприятия описывают законы цветного зрения, выведенные по результатам психофизических экспериментов. На основе этих законов за период более 200 лет было разработано несколько теорий цветного зрения. Однако, приблизительно в последние 35 лет появилась возможность непосредственно проверить эти теории методами электрофизиологии путем регистрации биоэлектрической активности одиночных фоторецепторов зрительной системы, а также расширить область взаимодействия и количество фоторецепторов сетчатки и мозга в цветовосприятии. Например, в 2007 году признано открытие 1991 года фоторецепторов ipRGC (не колбочки и не палочки, расположенных в ганглиозном слое на пути световых лучей оптического изображения в сетчатке глаза).

В связи с проведенными ранее исследованиями сетчатки при помощи рентгеноскопии (у приматов)[19] и проводимые в настоящее время исследования живых клеток и в том числе клеток сетчатки (колбочек, палочек и ipRGC) на базе нового микроскопа — Флуоресцентного микроскопа с разрешающей способностью 1-10 нм, на атомно-молекулярном уровне, появилась возможность получать новые данные исследований в цвете (это важно для оценки восприятия цвета экстерорецепторами, чувствительными к определённой длине волны спектральных зон S,M,L с данными стереоизображений на мониторе в заданном масштабе, а также новых методов измерения цвета (см. Измерение цвета (версия Миг)) (см. рис. 4а). Так уже проведены работы при исследовании цветного зрения у птиц на живых клетках сечений сетчаток (см. Цветное зрение у птиц (версия Миг)) в 2006—2008 годах[20]. Открытие третьего типа фоторецепторов ipRGC в сетчатке дало возможность более глубже взглянуть в тайны работы зрительной системы, уточнить детали процесса восприятия света и цвета. Также удалось связать работу экстерорецепторов и фоторецепторов сетчатки (палочек, колбочек, ipRGC), расположенных в разных слоях, функционирующих в биологической среде дифференцировано и в единой системе с участием коры головного мозга (осознанно).
В данной биологической системе функционирует прямая и обратная связь между экстрорецепторами и мозгом, при которой образование оптического изображения происходит через несколько этапов: создание, получения «первичного» оптического изображения в сетчатке (на фоторецепторном уровне — палочек, колбочек) с передачей сигналов от изображения предметных точек в головной мозг. Нейронные сети, в режиме принципа «оппонентности» (см. Теория оппонентного цветного зрения (версия Миг)) формируют уже в нашем сознании оптический образ — «изображение» (в зрительных отделах головного мозга). При этом оптические изображения формируются перекрёстно в правом и левом полушарии головного мозга соответственно от левого и правого глаза в стерео, то есть бинокулярном зрении. И в основе трёхкомпонентного принципа RGB — трихроматизм заложено:
- 1) Создание чёрно-белых оптических изображений в сетчатке «контурно» — (рецепторный уровень),
- 2) Создания цветных оптических изображений в мозгу — (нейронный уровень).
Важными открытиями являются восприятие света экстеро и фоторецепторами сетчатки глаза — это Цветное зрение: у приматов (и человека), у птиц, у рыб и др. (см. Труды доктора Р.Е.Марка и его лаборатории, Цветное зрение у птиц (версия Миг), Ретиномоторная реакция фоторецепторов сетчатки глаза). В настоящее время принято: цветное зрение осуществляеся тремя колбочками S,M,L при дневном освещении, зрение в условиях слабого освещения — не цветное (чёрно-белое), осуществляется палочками. (Более подробно см. ниже).
Принцип трихроматизма — основа цветного зрения[править | править код]
.jpg)



Теория многокомпонентного цветовосприятия исторически основана на многие гипотетические теории, предсказательная сила которых была невелика. Даже гениальные предположения учёных прошлого были в основном умозрительны, так как естественные науки прошлого не могли расширяться, так как не обладали в своё время необходимым инструментарием; не был сформирован понятийный аппарат, невозможно было провести многие критически важные эксперименты в силу недостаточной разрешающей способности оптико-механических приборов (например, микроскопов). Бурное развитие естественных наук можно отнести начиная с ХIХ столетия, когда специалисты в области биологии, химии и физики сделали огромный прорыв в естественных науках. (См.Участник:Alexandr/Современные концепции световосприятия и цветного зрения).

Треугольник Cb-Cg-Cr это только xy=(0,0), (0,1), (1,0), треугольник CIE xy— цветность пространства. Линия, соединяющая Cb и Cr это линия нулевой яркости на цветовом графике (alychne).
Обратите внимание, что спектральный локус проходит через rg=(0,0) на 435.8 нм, через rg=(0,1) в 546.1 нм и через rg=(1,0) при 700 нм. (По старой системе применялось цветовое пространство RGB и рассчитывалось без учёта линейной функции, а область полученной цветовой палитры была внутри треугольника с углами в точках 430 нм, 540 нм и 570 нм).
Значение же энергии точки (E) при rg=xy=(1/3,1/3) равное. (См. также XYZ (цветовая модель), Цветовые координаты)


На базе полученных новых данных максимумов спектральной чувствительности фотопигментов клеток сетчатки (S,M,L), палитра цветов, которую может различать глаз, работающий в соответствии с теорией трёхкомпонентного цветного зрения с учётом всех фоторецепторов сетчатки глаза и мозга, имеет вид согласно диаграммы палитры. На рис.2а,3а,Fig.15 видим области цветовой палитры, характеристику графиков кривых цветного восприятия колбочек с оппонентным отбором самых ярких основных, базовых цветов S,M,L (RGB, XYZ (цветовая модель), Цветовое пространство CIE 1931), которые анализируются или синтезируются при известных максимумах чувствительности фотопигментов йодопсина (см. рис.3, Fig. 15, рис.2a).
Клетки ipRGC, расположенные в сечатке, в зоне Ganglioniс layer и рефлекторно связанны с палочками и колбочками, имеют прямую и обратную сязь с мозгом. Они учавствуют в обработке информации, поступающей изначально при зрении, являясь фильтром УФ падающих лучей и взаимодействуют с фоторецепторами сетчатки (колбочки и палочки) и мозгом, так как они непосредственно своими синапсами связаны с ними. Содержа фотопигмент меланопсин (воспринимает и фильтрует фиолетово-синие лучи) фоторецепторы ipRGC автоматически участвуют в работе колбочек-S и зрительных отделов головного мозга в создании оптического изображения.
В итоге нормальное объяснение трихроматизму состоит в том, что у обычного человека сетчатка глаза содержит три разновидности «цветных» фоторецепторов, названных колбочками сетчатки глаза. В зависимости от сфокусированной на них предметной точки изображения они способны воспринимать видимые лучи предметной точкии и выделять из них три вида основных, базисных самых ярких световых лучей (S,M,L); с последующей передачей их в мозг. При этом участвуют независимые фоторецепторы ipRGC. На рецепторном уровне происходит выделение основных сигналов RGB (S,M,L — «синих», «зелёных», «красных»). Фоторецепторы ipRGC связаны с ними (рецепторный уровень) и участвуют в последующей трансдукции этих биосигналов (бесцветных) в головной мозг. В мозгу происходит окончательное формирование (нейронный уровень) цветного, стерео оптического изображения (в зрительных отделах головного мозга) — появления ощущения цвета.
Визуальное цветное зрение[править | править код]
Визуальное цветное зрение — основа современных теорий цветного зрения. Цветовое зрение происходит в зрительной системе, где инициируется поглощение света с помощью трех различных спектральных классов «шишек» — колбочек. Следовательно, цветовое видение описывается как трёхвариантное, на основе трихроматизма восприятие основных цветов или как восприятие, ощущение цвета. Первоначально психофизические исследования показали, что цвета могут быть настроены на использование трех различных систем (праймериз). В 1802 году, Томас Молодых предложил модель, по которой восприятие цвета может быть закодировано на восприятие цвета трёх основные фоторецепторов, но не на кодировании тысяч цветовых рецепторов для отдельных цветов.[22]
Спектральная чувствительность колбочек[править | править код]

1 — мембранные полудиски;
2 — митохондрия;
3 — ядро (эллипс с жировой каплей);
4 — синаптическая область;
5 — связующий отдел (перетяжка);
6 — наружный сегмент;
7 — внутренний сегмент;
8 — граница мембранной части;
9 — пигмент сократимых фибрилл.

Спектральная чувствительность колбочек может быть определена посредством нескольких методов. Два из этих метода включают изолирующую рецепторные (receptoral) ответы (Бейлор et al., 1984) с использованием вычисленных от цвета функции нормалей и дихроматизма или двухкомпонентности восприятия цвета (dichromats) (Смит и покорного, 1975; dichromat является предметом, в основе которого сетчатка имеет одну колбочку с фотопигментом в мембране (photopigment), достаточного для этого), микроспекрометрии (microspectrometry) (Bowmaker и Dartnall, 1980) или на основе отражения световых лучей — денситометрии (Раштон, 1963, 1966). Метод микроспектометрии предполагает выделение одной колбочки, пропуская свет через неё. Изменения в передаче различных длин волн может быть использовано для вычисления спектрального поглощения колбочкой или возможность определить изменения в электрическом ответе. Отражение денситометрии включает режиссуру, структуру света в сетчатке и определение изменения в поглощении как функция длины волны. Эти результаты впоследствии используются для расчета спектральных поглощений.
Было принято три класса колбочек в человеческой сетчатке глаза, которые были изолированы от указанных технологий. Эти три класса (колбочек) «шишек» представляют:
- Короткие волны (S): чувствительность к ним S-колбочки (S-конуса),
- Средние волны (M): чувствительность к ним M-колбочки (М-конуса),
- Длинные волны (L): чувствительность к ним L-колбочки (L-конуса).
У всех разная, но пересекающаяся спектральная чувствительность. Спектральная чувствительность S-колбочек с пиком, примерно, в 440 нм, М-колбочек — 545 нм и L-колбочек с пиком в 565 нм после исправлений при предварительной потери света сетчаткой. Однако различные методы измерения дали результаты с несколько различными вариантами максимального значения чувствительности (рис.1).[23]

Трёхцветная природа цвета[править | править код]

Трехцветная природа цветового зрения позволяет практически получить любой цвет, при смешивании трех основных цветов (RGB). При этом trichromacy цветного зрения является линейной функцией. Это означает, что колориметрические уравнения имеют свойства обыкновенного линейного уравнения. (См. Линейное уравнение цветного зрения и цветового пространства).
Цвет, соответствующий количеству Cs стимула S, может быть выражен как:
Cs(S) = C1(1i) + C2(2i) + C3(3i)
где li-3i есть три основных цвета (не обязательно монохроматического цвета спектра) и C1-C3 — трехстимульные значения.
Трехстимульные значения представляют собой количества каждого из трех первичных выборов, необходимых для достижения соответствия для цвета и яркости (рис. 2c). Они могут быть выражены в единицах светового потока или потока излучения, или даже произвольным масштабом, при условии, что произвольный масштаба передает относительные пропорции трёх количеств. (Это RGB).
Трихроматизм и компонентный характер цветного зрения[править | править код]
- (Данный материал создан совместно с Алекандровым и учтён при написании данной статьи.)
Понятие трихроматизма вводится для цветного зрения, что бы выразить связь причины появления цветного зрения, цвета с биологисеским инструментом, создающий это явление как цветное зрение. В природе все биофизические и биохимические процессы подчинены общим законам, которые мы стремимся раскрыть. Не случайно луч света диспергируется на монолучи (спектр (версия Миг)), которые участвуют в зрительном процессе. Однако как оказалось, спектр содержит основные и не основные лучи света. Основными лучами оказались три монолуча: красный, зелёный, синий — то есть RGB (аббревиатура английских слов Rred, Ggreen, Bblue — красный, зелёный, синий) — аддитивная цветовая модель, как правило описывающая способ синтеза цвета для цветовоспроизведения. Откуда весь набор биологических структур цветного зрения подчинён закону аддитивного синтеза цвета. Например, это три цвета реализуются колбочками. Собственно отсюда и названия трихроматизм, трёхкомпонентный принцип цветного зрения. Это случай совпадения триады лучей и триадой фоторецепторов, участвующих (колбочек) при цветном зрении.
| Обозначения основных цветов и фоторецепторов | Обозначение фотопигментов (разновидностей опсинов) | Диапазон | Пиковая длина волны |
|---|---|---|---|
| S-Палочки — максимальный фиолетово-синий; | (родопсин) | 450-530 нм | 498 нм |
| S-Колбочки, ганглиозные фоторецепторы ipRGC — синий; | β (разновидность йодопсина), меланопсин | 400-485 нм | 437 нм |
| M-Колбочки — зелёный;, желтовато-зелёный; сине-зелёный; (красный-мало); | γ (разновидность йодопсина) | 455–610 нм | 533 нм |
| L-Колбочки — красный, желтовато-красный; | ρ (разновидность йодопсина) | 480–650 нм | 564 нм |
При этом существуют одно, двух, трёх, четырёх и более компонентные структуры цветного зрения, но в любом случае (биологическая) система зрения — глаз в результате эволюции имеет в принципе одинаковые структуры для процесса зрения. Это оптическая система, сетчатка с фоторецепторами и зрительные отделы мозга
На сегодняшний день учёные не гипотетически, а на опытах доказали и получили изображения трёх, четырёх и более типов разных колбочек, которые при фокусировании на них лучей предметных точек изображения оппонентно выделяют самые яркие, базовые основные спектральные монохроматические лучи света RGB в виде биосигналов для посылки их в мозг. Т.о. такая теория на сегодняшний день является основной. В таблице ЗОНЫ ОСНОВНЫХ ЦВЕТОВ RGB определены спектры поглощения фоторецепторами сетчатки, их спектральные максимумы поглощения видимого излучения света — это S, M.,L, зоны основных цветов трихроматизма (номинальные, наиболее яркие спектральные области работы фоторецепторов сетчатки при цветном зрении).(См. также Ретиномоторная реакция фоторецепторов сетчатки глаза,[24]).
Колбочки и цветное зрение[править | править код]
| КОЛБОЧКИ | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||
| |||||||||||||||
Ко́лбочки или шишки — (англ. cй7one) клетки являются одним из трех типов фоторецепторов в сетчатке глаза млекопитающих (например, глаза человека). Они отвечают за цветовое зрение и лучше функционируют в сравнительно ярком свете, в отличие от палочек, которые работают лучше при тусклом свете. Колбочки являются ответственными за цвет, цветное зрение; колбочки функционируют лучше всего в относительно ярком свете c длиной волн более 498 нм, в противоположность клеткам фоторецепторов палочек, которые работают лучше в тусклом свете с длиной волн менее 498 нм. Колбочки плотно упакованы в ямке фовеа «централь», с диаметром 0.3 мм стержня — свободная зона с очень тонкими, плотно упакованными колбочками, которые быстро уменьшаются количественно к периферии сетчатки глаза.[26] Три типа колбочек, как правило отличаются тем, что каждая из трех типов содержат различные пигменты, определяющие цветовое ощущение (зрение), а именно: S-колбочки (синий цвет), M-колбочки (зелёный цвет) и L-колбочки (красный цвет). Каждая колбочка поэтому чувствительна к видимым длинам волн света, которые соответствуют длинам волн: коротковолновой, средней длине волны и длинной волне света RGB.[27]
Цветное зрение с точки зрения физики[править | править код]
В последнее время (2009‒2011) учёные физики, доктора Джеральд К. Хат[4] (см. Центральная ямка сетчатки глаза (версия Миг)), John A. Medeiros[28] (см. Работа мембраны колбочек и палочек как волновод) внесли новый подход в интерпретацию цветного зрения с точки зрения физики, которая не противоречит, и даже уточняет некоторые полученные выводы, данные известных учёных, предшественников — биологов, биофизиков, биохимиков.
Сейчас учёные рассматривают колбочку как клетку с внешней долей конусной мембраны, имеющей пространственную структуру, работающей по типу конусного волновода в среде прозрачного тела глаза. Это даёт возможность ей воспринимать сфокусированные лучи предметной точки оптического изображения в очерёдности, как работает конусный волновод в прозрачном теле сетчатки глаза, то есть в очерёдности (обратной): красные→зелёные→синие→оранжевые и т. д.. При этом идёт отбор основныех спектральных квантованных лучей (S,M.,L,) или RGB (cм. Основные цвета (версия Миг)), притом не в цвете. Эти отфильтрованные квантованные лучи в виде длин волн предметной точки со своими характеристиками преобразуются, трансформируется со скоростью в фемтосекуду, а затем трансдукцируются в мозг, где сфокусированная квантованная предмтная точка оптического изображения (в квантах энергии её длины волны) не в цвете, сканируется со своими координатами, но уже в цвете. Затем сигналы проводятся по нервным стволам в зрительные отделы головного мозга, где преобразовываются у нас в мозгу в виде аналогового оптического изображения в цвете, стерео изображение. Это бинокулярное оптическое изображение (субъективное), которое создаётся у каждого из нас своё.

Никакой заменитель глаза не способен это оптическое изображение повторить, мы можем с применением современной техники приблизиться к глазу, познать его принципы работы, но в настоящее время существует много непознанных вопросов получения оптического изображения на сетчатке, в мозгу. Самое важное, созданная техника не может работать как пространственный биологический фоторецептор — конусная внешняя доля мембраны колбочки. Предполагаемый принцип работы колбочки (её внешней конусной мембраны) в режиме конусного волновода помещённого в среду прозрачного тела глаза сводится также к принципу оппонентного отбор основных (RGB) квантованных спектральных лучей света не в цвете (рецепторный уровень). Например, самый совершенный цифровой фотоаппарат, колориметр, которые предназначены для получения цвета при воздействии на их фотодатчики, примерно как у глаза, на рецепторном уровне фиксируют оптическое изображение каждой предметной точки не цветное. Главное отличие состоит в том, что не цветное оптическое изображение в системе RAW (без цвета) и в оптических приборах воспринимается плоским фотоприёмником — фотосенсором или фототранзистором. Например, Фото-датчик, состоящий из мозаики ячеек трёх, четырёх пикселей, накрытых светофильтрами системы RGB (см. Рис. Б; Мозаика из ячеек фильтра Байера RGGB) содержит каждую клетку (блок) из «красного», «зелёного», «синего» пиксела (система RGB — трихроматизма), которая фиксирует сфокусированную предметную точку в виде трёх основных спектральных отфильтрованных лучей (трихроматизма) с квантованной энергией, равной длине волны, в виде заряда конденсатора-пикселя с определённым количеством электронов. Ещё с той разницей, что каждая сфокусированная предметная точка с полным набором её спектральных лучей оценивается тремя пикселями, несущие информацию трёх рядом расположенных сфокусированных предметных точек по одному поглощённому спектральному лучу (набора RGB. И каждая предметная точка, сфокусированная на пиксель, где каждый пиксель отбирает один основной спектральный луч из трёх падающих на него — или красный, или зелёный, или синий, фиксирует энергию длины волны одного из трёх лучей. (Так как каждый пиксель накрыт светофильтром красным, зелёным, синим). То есть 1/3 цветовой информации каждой сфокусированной предметной точки. Для создания цветного изображения, в результате полученных данных, применяется АЦП, который формирует из полученных сигналов цветное оптическое изображение, но не аналоговое. (См. Аналоговая фотография).
Как видим, биологическая система зрения при формировании оптического изображения на чёрно-белом и цветном уровнях у нас происходят в отличие от научно-технических достижений (цветная фотография) по законам биофизическим и биохимическим имеют общие сходства, но и принципиальные отличия от электронно-оптических систем.
- Главные отличия:
- 1) Оптическое изображение в нашем сознании — аналоговое, индивидуальное и для нас является эталоном, согласно которому мы сравниваем со всеми изображениями, которые нас окружают, или которые мы создаём сами.
- 2) Наш фотоприёмник — сетчатка глаза с жёлтым пятном с центральной ямкой, где формируется 50 % объектов цветного зрения и где расположены одни колбочки.
- 3) Главным фотодатчиком биологического фотоприёмника при цветном зрении в настоящее время является колбочка, которая с точки зрения чисто физической рассматривается как клетка с внешней долей конусной мембраны, имеющей пространственную структуру; и работающей по типу конусного волновода в среде прозрачного тела глаза. Это даёт возможность ей воспринимать сфокусированные лучи предметной точки оптического изображения в очерёдности согласно принципа работы конусного волновода, то есть очерёдности восприятия падающих на него спектральтных лучей в виде порядка красные→зелёные→синие→оранжевые и т. д., при этом оппонентно отбирать самые яркие основные, базисные спектральные лучи сфокусированной предметной точки (S,M,L,) или RGB не в цвете (внешняя мембрана — биологический конусный волновод внешней доли конусной мембраны колбочки). Если для получения цветного изображения фотокамерой применяется фотосенсор, где ячейка из пикселей RGB фильтрует только одну составляющую цвета сфокусированной предметной точки, то есть 1/3, то для получения всех трёх цветов при помощи АЦП из трёх клеток фотосенсора (трёх предметных точек в каждой, рядом расположенных) формируется цветное изображение не аналоговое, но ещё более красочное, чем оригинал. В мозгу же, на основании отобранных сигналов более коротким путём, при этом аналоговых сигналов каждой предметной точки, формируется полноценное наше аналоговое цветное оптическое изображение.
- 4)Зрительные пигменты колбочек (кон-опсины) сконцентрированы в наружных дисках мембраны. В колбочках пигмент расположен в особых пигментных складках, которые являются продолжением наружной клеточной мембраны экстерорецептора. Молекулы пигмента составляют около 80 % всех белков диска. Зрительные пигменты настолько плотно упакованы в мембранах наружного сегмента, что расстояние между двумя молекулами зрительного пигмента не превышает 10 нм. Такая плотная упаковка повышает вероятность того, что фотон света при прохождении слоя фоторецепторных клеток будет уловлен. (Например, каждая палочка содержит около 108 молекул пигмента. Они организованы в несколько сотен дискретных дисков (около 750 в палочке обезьян), которые не связаны с наружной мембраной.)
- Каждая колбочка содержит фотопигменты кон-опсины (Йодопсин (версия Миг)) : красные, зелёные, синие, которые участвуют в оппонентном выделении из луча одного из трёх основных, базовых биосигналов S,M,L не в цвете. При оппонентном отборе биосигнала в мембране колбочки происходит сравнение трёх пар сигналов красный-зелёный, синий-жёлтый, чёрный-белый. Оппонентно отобранный биосигнал отправляется по нервным стволам в зрительные отделы головного мозга.[29],[30] (См. также Теория оппонентного цветного зрения).
Отличающиеся особенности колбочек от палочек[править | править код]
1 — мембранные полудиски;
2 — митохондрия;
3 — ядро (эллипс с жировой каплей);
4 — синаптическая область;
5 — связующий отдел (перетяжка);
6 — наружный сегмент;
7 — внутренний сегмент;
8 — граница мембранной части;
9 — пигмент сократимых фибрилл.


Отличающиеся особенности колбочек и палочек состоят в том, что колбочки (см. рис.1а) морфологически однотипны, но в зависимости от места расположения на сетчатке имеют отличающиеся размеры. у них у всех внешняя мембрана конусообразная, но отличается в зависимости от места расположения. Это связано с функцией их работы. Но их три и все они в принципе созданы для восприятия трёх основных лучей RGB с большим перекрытием диапазона волн. Например, колбочки в центральной ямке имеют большую плотность расположения и воспринимают в основном красно-зелёные лучи, они имеют больший размер основания конуса внешней доли мембраны, так как поперечное сечения фронтального луча красного цвета наибольшее и соответственно основание конуса мембраны большее, чем у зелёной, а тем более синей колбочки, которая находится вне центральной ямки фовеа. (Работа мембраны колбочек и палочек как волновод). Все колбочки содержат разновидности фотопигмента кон-опсины, которые в зависимости от захваченного луча света предметной точки и оппонентного отбора выделения наиболее яркого основного луча триады RGB (S,M,L) (cм. Основные цвета (версия Миг)) в мембране выделяется фотопигмент кон-опсин данного цвета. У колбочек три разновидности фотопигмента йодопсина — фотопигменты хлоролаб и эритролаб. Фотопигмент хлоролаб поглощает лучи жёлто-зелёной части спектра света. Фотопигмент эритролаб поглощает лучи жёлто-красной части спектра. Третий вид фотопигмента цианолаб (версия Миг) (предполагаемый) поглощает синие лучи света, который проявляется при захвате синих лучей колбочкой-S. Колбочки-S располагаются вне зоны ямки, в периферийной зоне сетчатки.
Однако пока известно, что Йодопсин (версия Миг) находится во всех колбочках глаза.
Отличающаяся особенность палочек (см. рис.2b) состоит в том, что палочки имеют мембрану цилиндрической формы с поперечным сечением порядка 1,5-2,5 мкм, что соответствует поперечному сечению фронта волны синих лучей. Палочки отсутствуют в центральной ямке и работают в условиях сумеречного и ночного освещения.
Плотность и распределение колбочек и палочек в сетчатке[править | править код]
.jpg)
Для понимания организации визуальных связей важно знать пространственное распределение различных типов клеток в сетчатке. Известно, что фоторецепторы организованы с довольно точной мозаикой. Как мы видели в фовеа, мозаика представляет собой гексагональную (шестигранную) упаковку колбочек. За пределами ямки палочки разрушают закрытую гексагональную упаковку колбочек, но при этом позволяют организованную архитектуру с колбочками, равномерно распределенной системой палочек (восемь или шесть) вокруг окружностей палочек. Таким образом, в терминах плотностей различных популяций фоторецепторов в сетчатке человека ясно, что плотность колбочек самая высокая в ямке фовеа и быстро падает за пределы ямки до довольно ровной плотности в периферической сетчатке (фиг.20 и 21) (Osterberg, 1935; Curcio et al., 1987).[33][34] Существует пик палочковых фоторецепторов в кольце вокруг ямки около 4,5 мм или 18 градусов от фовеальной ямы. Оптический нерв (слепое пятно), конечно, свободен от фоторецептора (см. Ниже).[35]

Распределение колбочек и палочек в сетчатке — когда в поле зрения человека входит большой набор центральных и не центральных точек сетчатки глаза, важно знать пространственное распределение различных типов клеток в сетчатке для понимания организации визуальных связей и их строение.
Точки, входящие в понятие центральное зрение — центральная ямка сетчатки глаза (версия Миг), а понятие не центральное зрение — периферийное зрение.
- Центральное зрение (5°) (см. рис. 3).
- Периферийное зрение разделяется на три пояса (см. рис.4):
- 1) Ближне-периферийное иногда называемое Пара-центральное зрение, находящееся по соседству с центром взгляда. [нужная цитата]. Именно область Пара-центральное зрение содержит наибольшее количество синих S-колбочек, которые участвуют в оппонентном отборе основных базовых лучей кружка нерезкости RGB сфокусированной предметной точки в зоне центральной ямки с колбочками L,M. сетчатки.
- 2) Средне-периферийное зрение, находящееся в области середины поля зрения (mid perip.); Пара-центральное зрение,
- 3) Далеко-периферийное зрение, находящееся в области, расположенной на краях поля зрения (far perip.);[37]
Ещё в 1970-х годах было показано, что распределение типов колбочек в сетчатке неравномерно: «синие» колбочки находятся ближе к периферии, в то время как «красные» и «зеленые» распределены случайным образом. Однако, Мы описываем подробный анализ пространственного расположения конусов L, M и S в живых глазах двух людей и одной обезьяны. Мы анализируем мозаику конуса около 1 градуса эксцентриситета, используя статистические методы, которые характеризуют расположение каждого типа конуса в мозаике фоторецепторов. Во всех глазах конусы M и L расположены случайным образом. (Мозаика в центре, центральной ямки фовеа — расположены только колбочки M/L c шестиугольной упаковкой (см. рис. P/T)). Это приводит к появлению патчей (исправлений, заплаток), содержащих конусы одного типа. В человеке, но не в обезьяне, расположение S-конусов нельзя отличить от случайных.[38] Т.е. распределение колбочек и палочек доказанное Robertом E. Marcом [39] не опровержимо.
Своеобразие работы синих S-Колбочек[править | править код]

Своеобразие работы синих S-Колбочек отличается от красных и зелёных своим местом расположения (мозаикой) в фокальной поверхности сетчатки лаза, а также их работой — восприятием синих лучей видимого спектра света в отличии от работы красных и зелёных колбочек. (См. рис.1,1p,2). Например, в центральной ямке фовеа синие колбочки-S насчитывают 12 % числа колбочек L,M, остальные расположены в периферийной зоне центральной ямки с изменяющимся размерами и уменьшением плотности расположения по мере удаления от границы периферии.
- За последние несколько лет психофизики, электрофизиологи, генетики и анатомы пришли к выводу, что в коротковолновой системе есть что-то уникальное по сравнению с двумя более длинноволновыми системами в визуальной системе. [16]
.jpg)
Из трех спектральных типов колбочек RGB, обнаруженных в нормальной сетчатке человека, только одну S-колбочку или синюю колбочку можно отличить от других в мозаике. Используя специальные антитела, генерируемые против колбочек с разновидностью синего пигмента опсина, являющиеся визуальными пигментами, содержащиеся в колбочках, можно селективно окрашивать коротковолновые чувствительные пигментные (или синие пигментные) S-колбочки. (рис.3) (Szell et al., 1988; Ahnelt and Kolb, 2000).

Существуют различия в генетической структуре и локусе (Локус (лат. locus — место) в генетике означает местоположение определённого гена на генетической или цитологической карте хромосомы.) визуального пигмента S-колбочеи по сравнению с пигментами L- и M-колбочек (Nathans et al., 1986), однако S-конусы являются общими для всех позвонков позвоночных и всегда образуют своё место в пермферийной зоне (Marc, 1982, Kolb and Lipetz, 1991). Тем не менее S-колбочек очень мало в центре фовеа, что вызывает так называемое слепое пятно S-колбочки (Williams et al. ., 1981), и их минимальное количество в центре фовеального склона — около 12 % числа всех колбочек фовеа. (рис.2).
Дифференцированная работа экстерорецепторов[править | править код]
На основании последних данных (См. рис. 1,1а,1г, доктор Джеральд К. Хат[42] и доктор John A. Medeiros[2]учёные вообще, хоть и немноо гипотетически, на базе всего набора информации по цветному зрению, более реалистично пришли к обобщению, что цветное зрение следует рассматривать с точки зрения дифференцированной работы на фокальной поверхности сетчатки фотодатчиков в виде нано-антен или структур колбочка в окружении шести или восьми палочек. Каждая нано-атена — пространственная структура, что согласуется с тезисом, что каждая колбочка работает как конусный волновод в среде прозрачного тела глаза. Данное направление снимает вопросы хроматической аберрации, фильтрации основных лучей, наличия необходимых пигментов, определения в сетчатке только трёх, геометрически-определенных длин волны и обнаружение среди них максимальных границ, исключительно длин волн: от длинной волны до короткой (700нм и 400нм) визуальной полосы и, даёт критически точное, геометрически определённую опорную, базовую (midband) длину волны (550 нм) для нахождения величины длины волны синего цвета (S).
Например, подтверждая вышесказанное, в работе Helgе Kolbа дано:
- Электронная микроскопия, наконец, показала, что тип HII горизонтальной клетки действительно послал много древовидных «процессов» (сигналов) немногим синим колбочкам (колбочки-S) посредством её древовидных небольших и меньших концентраций процессов, приведших к лежанию «M.» и L-колбочек (зелёных и красных). Короткие аксоны этих клеток HII связываются с конусами исключительно как на рис. 8b,9 (Ahnelt и Kolb, 1994). Внутриклеточная регистрация от горизонтальных клеток H2 в сетчатке обезьяны доказала окончательно, что эта горизонтальная синяя клетка (синяя колбочка) — чувствительный и важный элемент тропы колбочки в сетчатке примата (Dacey и др., 1996).[43]
Процесс фототрансдукции[править | править код]
Процесс фототрансдукции — важный и сложный процесс, объясняющий механизм поглощения фотонов, преобразования и передачи электрических биосигналов фотодатчиками. И чтобы понять его, нужно иметь представление о структуре клеток фоторецепторов, вовлеченных в зрительный процесс: о палочках, колбочках и клеток внутренних слоёв сетчатки. Клетки колбочек и палочек содержат хромофор, (11-cis-retinal, Альдегид Витамина A1[44] и легко-абсорбирующей части) обязанный белку мембраны клетки опсину (opsin). Палочки имеют дело со слабым освещением (низким сумеречным уровнем) и не добиваются цветного видения. Колбочки, с другой стороны, могут закодировать цвет изображения через сравнение продукций трёх различных типов колбочек! Каждый тип колбочки лучше всего отвечает на определенные длины волны, или цвета, света, потому что каждый тип имеет разновидности фотопигмента опсина (opsin). Три типа фоторецепторов: L-колбочки, М-колбочки и Булочки (S-колбочки) оптимально отвечают на длинные длины волны (красноватый цвет), средние длины волны (зеленоватый цвет), и короткие длины волны (синеватый цвет) соответственно.[44][45] Амакриновые клетки играют важную роль в вопросах регулирования и передачи сигналов колбочек и палочек в мозг. Они образуют чрезвычайно сложную, соединенную щелевыми контактами сеть, а отросток, не покидая пределов сетчатки, ветвится в области синаптических контактов биполярных ганглиозных нейронов, которые изменяют характер распространения возбуждения сигналов по нервным клеткам ганглиозного слоя.
Фоторецепторы и фотопигменты сетчатки[править | править код]
| Фоторецепторные клетки Колбочки, Палочки и ганглиозные клетки ipRGC
(Свет снизу) | |
|---|---|
 Рис. Ф.[46]
Стрелка и пунткирна линия — внешняя пограничная мембрана | |

Фоторецепторные клетки сетчатки глаз — расположенные в сетчтаке глаза (Экстерорецептры Колбочки (сетчатка) (версия Миг) и Палочки (сетчатка) (версия Миг) и фоторецепторы Фоторецепторные клетки ганглиозного слоя сетчатки ipRGC), воспринимают электромагнитное излучение в световом диапазоне длин волн. Фоторецепторы обеспечивают и создание оптического изображения — функцию зрения, и внешнюю коррекцию биоритмов сна и бодрствования, зависящую от общей освещённости.
Это группа специализированных светочувствительных образований в сетчатке, воспринимающих свет и по-разному возбуждающихся от воздейстаия них световых лучей, с ответом в виде фототрансдукции (передачи «преобразованного» сигнала). Фоторецепторы создают т. н. потенциалы действия — нервные импульсы, дополнительно обрабатываемые в зрительной коре головного мозга, при помощи совокупного движения, поведения биологических систем, которые замкнуты и одновременно имеющие модулятор движения (рефлексы), связывающий фоторецепторы и всех его внутренних элементов с движением внешней среды (электромагнитнх волн), из которой поступает движущийся биосигнал, и регулирующее по амплитуде или частоте движение саморегуляции биологических систем. Например, регулировка функции фочувствительного элемента фоторецепторов (мембран колбочек, палочек) означает автоматическую поднастройку оптимального положения их при раздражении падающими фокусирующимся основными спектральными лучами RGB. При этом диспергированный «белый» луч света должен иметь как минимум семь спектральных лучей фокусировки спектра вдоль оси мембраны колбочек на разной высоте, расположенных в блоках. Каждый блок имеет диаметр примерно 7-9 мкм (совпадающий с площадью, сфокусированного блока двух предметных точек с просветом на расстоянии 250 мм размером 0,176мм — разрешаюшая способность нормальной зрительной системы человека с остротой зрения 1,0 (см. Острота зрения человека)). Каждый сфокусированный луч предметной точки фокусируется в зоне конуса мембраны с диаметром основания в 3‒5мкм и высотой = 50мкм. В блоке может разместиться, примерно, более 3 колбочек, что достаточно с точки зрения теории оппонентности Эвальда Геринга — обработки цветового воздействия. То есть сфокусированные лучи любой предметной точки изображения на колбочки в виде кружков нерезкости (например, при остроте зрения 1.0 — разрешающая способность глаза 0,176мм) диаметром ~7-8 мкм каждой колбочкой оппонентно выделяется из пучка монолучей один самый яркий основной, базисный биосигнал не в цвете (красный, зелёный, синий) из трёх S,M,L который отправляется в мозг.[48][49] (См. также Теория оппонентного цветного зрения (версия Миг)).
Замечание[править | править код]
При рассмотрении вопросов визуального цветного зрения следует различать и отличать понятия яркость света (версия Миг) — физическая величина от яркости цвета — биологическая величина.
Яркость цвета связана с цветным зрением, нашим личным, биологическим восприятием световых видимых более слабых лучей (электромагнитных колебаний) (см. дневное зрение (версия Миг)). Так с колбочками S,M,L, (синих, зелёных, красных) с пиком длиной волны более 496 нм, которые нашим глазом воспринимаются как очень яркие (вопросы приспосабливаемости и выживания живых организмомв), хотя они физически по энергетике более слабые. У них частота колебаний волн более низкая, чем у синих, УФ лучей (длина волн менее 498нм). Дневной образ жизни животных связан с окружающей средой обитания, где в основном все объекты освещены дневными лучами света, а прямой и отражённый видимый спектр света содержит основные видимые лучи S,M,L,, которые более слабые, но биологически отбираются как наиболее яркие. Понятно, почему мы не видим Уф лучи, рентгеновские лучи и т. д. Природа выбрала свой вариант восприятия среды обитания и защиты глаза от ненужных ей сильных УФ, фиолетовых, высокочастотных синих лучей с длинами волн менее 498 нм. Например, синие, УФ лучи с длинами волн менее 496 нм для глаза являются не яркими, и колбочками не воспринимаются, так как они блокируются от попадания на колбочки ганглиозными и биполярными клетками сетчатки глаза, хотя они более мощные! (Парадокс). (См. рис. Ф).
Для восприятия лучей света при слабом освещении в условиях сумеречного и ночного зрения — «монохромных лучей» с длинами волн менее 498нм, в условиях «ночного видения» служат экстерорецепторы, называемые палочками, которые имеют пик чувствительности вокруг 498 нм и менее с фотопигментом высокой чувствительности при слабом освещении родопсином к лучам синим и УФ с высокой частотой колебаний (менее 496нм). (Колбочки их не воспринимают).
Откуда биологические понятия яркости и контрастности цвета при зрении отличаются от физических понятий яркости и контрастности света.
Эволюция и генетика фотопигментов[править | править код]
Впервые существование колбочкого пигмента (косвенным образом) было обнаружено Уолдом, который и дал ему название йодопсин.[50] В 1967 г. за эти работы ему была присуждена Нобелевская премия по физиологии и медицине.

Эволюция и генетика фотопигментов — направление исследований об эволюции и генетике фотопигментов фоторецепторов, связанной с наличием аутосомальных[52]генов, которые производят фотопигменты: в клетках колбочек сетчатки глаза (S-пигменты, «синие» колбочки), и X-связанные пигменты (см. рис. Э,Ф,ф/1) с пиковой чувствительностью в зелёном и красном диапазоне длин волн (M/L-пигменты, «зелёные» и «красные» колбочки).
Йодопсин и цианолаб[править | править код]
Йодопсин — один из зрительных пигментов опсинов колбочек человека и животных, точнее — группа пигментов. В настоящее время установлено, что это группа изоморфных пигментов, несколько различающихся по составу и спектрам поглощения.
У человека идентифицировано три вида иодопсина (фотопсина):
- 560 nm: rot-sensitives L-Iodopsin mit L-Photopsin, Шаблон:UniProt. Defekte am OPN1LW-Gen sind die Ursache der Protanopie
- 530 nm: grün-sensitives M-Iodopsin mit M-Photopsin, Шаблон:UniProt. Defekte am OPN1MW-Gen sind die Ursache der Deuteranopie
- 420 nm: blau-sensitives S-Iodopsin mit S-Photopsin, Шаблон:UniProt. Defekte am OPN1SW-Gen sind die Ursache der Tritanopie
Получены снимки колбочек (см.рис. S), графики трёх колбочек (см.рис.1), воспринимающих основные лучи RGB — S,M,L, на которых видны графика точек синих лучей S колбочки-S, которые она выделяет, видны сами синие колбочки, снятые в плане фокальной поверхности сетчатки. Т.е вопрос не открытого фотопигмента йодопсина цианолаба не означает, что нет синей колбочки, и что разновидности опсина не содержат пока не открытый синий фотопигмент опсинов — ЦИАНОЛАБ, который находится в колбочках.
Фоторецепторные клетки сетчатки глаза[править | править код]
.jpg)
Фоторецепторы ipRGC[править | править код]
.jpg)
Третий вид фоточувствительных клеток (см. Сечение сетчатки глаза) (не колбочки и не палочки), которые относятся к сетчатке глаза[54].
- pRGC (Retinal ganglion cell) — все фоточувствительные нервные клетки;
- ipRGC — фоточувствительные нервные клетки D,E, относящиеся к сетчатке глаза. (Фоточувствительные клетки нервного узла ipRGC содержат фотопигмент меланопсин[55] и расположены в сетчатке глаза не в фокальной поверхности сетчатки, где расположены экстерорецепторы колбочки и палочки, а — в ганглиозном слое сетчатки (Ganglionic layer) (см. Сечение сетчатки глаза).)
Клетки нервного узла (ipRGC), содержат пигмент (melanopsin)[56], — третий вид фоторецепторов сетчатки глаза недавно обнаруженый и являются типом нервной клетки в сетчатке[57] глаза млекопитающих. В отличие от других нервных узлов[58] сетчатки глаза, они являются фотосенсибилизационными[59]!. Это означает, что они — третий класс фоторецепторов сетчатки глаза, которые возбуждаются под действием лучей света. (Они даже при блокировании классических фоторецепторов палочек и колбочек, или в случае применения фармакологических агентов их блокировки, или при отделении их от сетчатки, независимо функционируют), т.е. участвуют в зрительном процессе.
Палочки[править | править код]

Па́лочки англ. rod cells) — один из типов экстерорецепторов, расположенных в фокальной поверхности сетчатки в слое (экстерорецепторов), периферических отростков светочувствительных клеток сетчатки глаза. Назван так за свою цилиндрическую форму. Это высокоспециализированные клетки, преобразующие световые раздражения в нервное возбуждение (сигналы).
Палочки чувствительны к свету благодаря наличию в них специфического пигмента — родопсина (или зрительный пурпур). Палочки работают в условиях сумеречного и ночного освещения.
Бесцветные рецепторы вечернего видения (scotopic видение) — палочки, так называемые из-за их формы. (См. рис. 1r).
Колбочки[править | править код]
Ко́лбочки (см.рис.1) — (англ. cone) один из типов фоторецепторов (Экстерорецепторы фокальной поверхности сетчатки глаза), расположенных в фокальной поверхности сетчатки в слое (экстерорецепторов), периферических отростков светочувствительных клеток сетчатки глаза, названный так за свою коническую форму. Это высокоспециализированные клетки, воспринимающие и преобразующие световые сигналы (фотоны) в нервное возбуждение в виде биоэлектрических (сигналов) на рецепторном уровне (есть уровень нейронный — участие в работе зрительных отделов головного мозга) на базе принципа оппонентного отбора более ярких сигналов RGB.
Колбочки чувствительны к свету благодаря наличию в них специфического пигмента йодопсина — разновидности пигмента кон-опсина. Колбочки работают в условиях дневного света, обеспечивая цветное зрение.

Также известно, что форма колбочки не идентична всюду в человеческой сетчатке (см. рис. 1с). Фоточувствительная внешняя часть доли мембраны колбочек изменяется систематически, чтобы быть длинной и мягко сужающимся, образованием конусности в центральной (foveal) части сетчатки к тому, чтобы быть более короткой, и более приземистой (более очевидно сужающейся, с большей конусностью) в периферийных частях сетчатки (von рисунки Грефа морфологии конуса по различным областям сетчатки показываются ниже). Coincidently, цветная функция видения, обеспеченная конусными внешними долями мембраны колбочки, также изменяется систематически в разных частях сетчатки от центра до периферийных. Цветное зрение имеет лучшее решение в зависимости от конусности мембраны (трехмерный или trichromaticy), и там, где конусы долго и мягко сужаются, c уменьшением в решении конусности (чтобы быть чрезвычайно двумерным или двуцветным) в промежуточных областях сетчатки, и уменьшается далее к ее самому низкому решению конусности (быть чрезвычайно одномерным или монохроматическим) в далекой периферии сетчатки (хотя цветная функция видения полностью не потеряна для достаточно ярких или больших участков цвета).
Фотохимические реакции в фоторецепторных клетках сетчатки глаза[править | править код]
Фотохимические реакции — химические реакции, которые инициируются воздействием электромагнитных волн, в частности — свет (версия Миг)ом. Примерами фотохимических реакций являются фотохимические реакции, которые инициируются воздействием электромагнитных волн, в частности — светом на фоторецепторные клетки сетчатки глаза, фотосинтез в растениях, распад бромида серебра в светочувствительном слое фотопластинки, превращение молекул кислорода в озон в верхних слоях атмосферы, фотоизомеризация, фотохимически инициируемые перициклические реакции, фотохимические перегруппировки (напр. ди-π-метановая перегруппировка) и т. п.
Ретиномоторная реакция фоторецепторов[править | править код]

Слева — микрофотография сечения сетчатки глаза рыбы, справа — схематическая прорисовка взаиморасположения органелл и экстерорецепторов. Направление света, приходящего в сетчатку сквозь хрусталик: снизу-вверх.
На фото видно: днём палочки скрыты гранулами меланина; рецепторная часть колбочек выдвинута к свету (на фото — смещена вниз).[62]

Справа — схема перемещений органелл и изменения формы фоторецепторных клеток.
На фото видно: при недостатке света гранулы меланина открыли путь света к палочкам; рецепторная часть колбочек удалена от источника света, рецепторная часть палочек — приближена к нему (на фото — смещена вниз).[63]
Ретиномоторная реакция фоторецепторов сетчатки глаза — механические процессы в сетчатке глаза, связанные с перестройкой взаимного расположения экстерорецепторов (палочек) и (колбочек), и гранул меланина,
в соответствии с уровнем освещённости (см. рис. 1/1,2/2,C), где видно, что колбочки работают при дневном освещении без участия палочек, а палочки работают при сумеречном и ночном освещении без колбочек! (Это важное открытие).
Цветное зрение у человека и приматов[править | править код]
Механизмы трехцветного световосприятия у приматов (Trichromatic) лучше всего исследованы на примере человека. Согласно последним достижениям в исследованиях физиологии зрительной системы животного мира, у остальных приматов зрение (версия Миг) незначительно отличается от восприятия цвета человеком.
Следует заметить, что у части людей, вследствие определённой мутации, возник дополнительный зрительный пигмент, с несколько иным строением и иной спектральной чувствительностью. У таких людей (а пока известно, что эта мутация передаётся только женщинам) зрение четырёхкомпонентно, то есть они несколько лучше различают оттенки различных цветов.
Цветное зрение у людей и животных связано с вопросами саморегуляции организма, с наличием совокупного движения, поведения биологических систем, которые должны быть замкнутыми и одновременно должны иметь модулятор движения, связывающий организм и его внутренние системы с подвижностью, колебаниями внешней среды, из которой поступают сигналы, воздействующие на элементы нервных узлов. Например, ретиномоторная реакция фоторецепторов — механические процессы в сетчатке глаза, связанные с перестройкой взаимного расположения экстерорецепторов (палочек и колбочек), и гранул меланина,
в соответствии с уровнем освещённости (см. рис. 1/1‒2/2).
Этот процесс, по сравнению со скоростью движения хрусталика, или реакцией глаза на движение, довольно медленный. Скорость адаптации легко оценить по времени, которое требуется нашим глазам для привыкания к резкой смене освещения (например, при переходе из солнечной комнаты — в тёмную, и наоборот, мы временно, на секунды, «слепнем»); но полная световая адаптация занимает 10‒30 минут. Такая саморегуляция функций организма — подстройка чувствительности органа зрение (версия Миг) к условиям освещённости, связана и с организацией циркадных ритмов[64] и обеспечивает адаптацию глаза к различным условиям окружающей среды, и переход от дневного, цветного зрения — к ночному, более светочувствительному, но монохроматическому. Наиболее ранние, а затем и самые подробные работы по изучению процесса ретиномоторной адаптации были проведены на рыбах.[65]
Физиология цветного восприятия[править | править код]

Восприятие цвета начинается со специализированных клеток сетчатки глаза, содержащих Фотопигмент (версия Миг) опсины (версия Миг) с различной спектральной чувствительностью, известных как колбочки сетчатки глаза. В организме человека существует три типа колбочек, чувствительных к трём различным спектрам, в результате чего у нас трехцветное цветовое зрение.
Каждая отдельная колбочка содержит зрительные пигменты (опсины (версия Миг)), состоящие из апротеинов (на базе G-белков), ковалентно связанных либо с 11-цис-hydroretinal или, реже-11-цис-dehydroretinal.[67].
Колбочки, условно названные по порядку длин волн из пиков их спектральной чувствительности (см. рис.1b):
- типы колбочек с короткой длиной волны (S),
- со средней длиной волны (М),
- с длиной длиной волны (L).
Эти три типа колбочек не соответствуют хорошо определенным цветам, как мы их знаем. Скорее, восприятие цветов достигается комплексным процессом, который начинается с дифференциальной работой этих клеток в сетчатке, и она будет завершена в зрительной коре и ассоциативных зонах зрительной коры головного мозга.
Например, в то время как L колбочки называли просто красными экстерорецепторами, microspectrophotometry Денситометрия (версия Миг) показала, что их пик чувствительности в зеленовато-желтой области спектра. Аналогичным образом, S и M колбочки напрямую не соответствуют синим и зеленым, хотя они часто изображаются как таковые. Важно отметить, что RGB — это всего лишь удобное средством для представления цвета, и не имеет прямой зависимости от типа колбочек в глазу человека.
Пик реакции человеческой колбочковой клетки меняется, даже среди лиц с «нормальным» цветовым зрением;[68] у некоторых видов, кроме человека, это полиморфные вариации — больше того, это вполне может быть адаптивным.[69],[70]. Например, явление метамерии, когда разные цвета человек ощущает как один цвет.
Трихроматизм и принцип оппонентности[править | править код]

Трихроматизм и принцип оппонентности — теория, которая определяет способ, которым сетчатка (версия Миг) человека (глаз (версия Миг)) обнаруживает и выделяет базовые лучи видимого света RGB с тремя разновидностями колбочек (S,M.,L) с использованием принципа оппонентного восприятия цвета (белый-чёрный, красный-зелёный, синий-жёлтый),[71][72]
Это происходит на уровне восприятия видимых лучей (рецепторном) тремя разновидностями колбочек RGB (S,M.,L), содержащих разновидности фотопигмента опсины (версия Миг) на базе оппонентного отбора цвета из цветов (красный-зелёный, синий-жёлтый (или синий-красный/зелёный)) с последующей передачей сигналов в мозг (нейронный уровень) (См. рис.9).
Цветное зрение у птиц[править | править код]
Доказано[3] (2006 год), что сетчатка (версия Миг) у птиц обладает одной из самых сложных систем фоторецептора колбочки среди позвоночных животных. Исследуя спектры поглощения сетчатки было получено, что птицы имеют пять типов колбочек, включая четыре одиночных, которые поддерживают видение цвета четырехроматизма(tetrachromatic) и двойную (колбочку), которая обеспечивает восприятие движения. (См. рис. 2a).
Когда пигмент поглощает свет (версия Миг), падающий на сетчатку глаза (на палочки и колбочки), изменяется мембранный потенциал клетки-колбочки, затрагивающий нейроны в слое ганглий сетчатки. Каждый нейрон в слое нервного узла может воспринимать информацию процессов от множества фоторецепторов, и может в свою очередь вызвать нервный импульс и передать информацию по оптическому нерву для дальнейшей обработки в специализированные зрительные центры мозга. Чем более интенсивный свет, тем больше фотонов поглощено зрительными пигментами, и большее возбуждение каждой клетки стимулирует появление более яркого оптического изображения.[3] При этом предварительно монохроматические диспергированные лучи перед попаданием в мембрану попадают в жировые капелки, где оппонентно фильтруются наиболее сильные, яркие сигналы, которые в результаце фототрансдукции фильтрутся (согласно их цветовой специализации) и направляются по зрительным нервным стволам в мозг. С учётом оппонентного принципа отбора более яркого сигнала на этапе рецепторной работы зрительной системы выделение сигнала производится с участием ганглиозных слоев нейронов сетчатки, параллельно с выделением занимаются подавлением второстепенных, слабых сигналов, сжатием потоков из 6 млн до 1,2 млн для передачи по 1,2 млн нервных каналов. При этом с участием зрительных отделов головного мозга, связанного с ганглиозными фоторецепторами, происходит фильтрация трансдукцируемых цветных сигналов через фоторецептор ганглиозных клеток сетчатки ipRGC в прохождении фиолетово-синих лучей благодаря наличию у них фотопигмента меланопсина.
К тому же цвет жировых капелек каждой колбочки, ещё до попадания сигнала в мембрану, (например, синий, зелёный, красный) яркое доказательство трихроматизма цветного зрения. То есть на этапе уже рецепторном восприятие цвета происходит колбочками с разновидностью фотопигмента кон-опсина, которые отбирают свои яркие спектральные области монолучей (S,M.,L) предметных точек оптического изображения.
Роль жировых капелек колбочек и палочек[править | править код]

В глазу многих видов рептилий, птиц и других животных имеются фоторецепторы — колбочки особого вида — с «жировой капелькой» в рецепторной части. Считается, что такие капельки, окрашенные липофильными природными пигментами — каротиноидами, выполняют роль предварительного, «внутриклеточного» светофильтра. Эти светофильтры дискриминируют из общего светового потока часть длин волн света, тем самым обеспечивая улучшенное цветоразличение.[74]
У таких животных нередко наблюдается 4-6 типов колбочек, и колбочки разного типа имеют жировые капельки с различной концентрацией каротиноидов (то есть имеют разные типы «встроенных светофильтров»). Предварительно отфильтрованный ими свет создаёт нервный сигнал разного типа благодаря восприятию соответствующим видом колбочек отдельных диапазонов цветового спектра. Затем по нервным клеткам через синапс сигнал попадает в зрительные нервные стволы, ведущие эти сигналы в мозг. Жировые капельки, окрашенные в цвета четыре-, пять- и более оттенков — яркое доказательство многоэтапности восприятия света и цвета сложными и разнообразными зрительными системами животных.[75]
Головной мозг и оптическое изображение[править | править код]

Зрительная кора является частью коры больших полушарий головного мозга, отвечающая за обработку зрительной информации. Главным образом она сосредоточена в затылочной доле каждого из полушарий головного мозга[76].
Оппонентно выделенные самые яркие сигналы видимых лучей света S,M,L — RGB (не в цвете), сфокусированных предметных точек на экстерорецепторы колбочки сетчатки глаза (рецепторный уровень), по зрительным нервам пересылаются сюда, в зрительную кору. Здесь формируется бинокулярное (стерео) цветное оптическое изображение (нейронный уровень). Впервые, субъективно мы ощущаем цвет (версия Миг), который является лично нашим. (При определении цвета методом колориметрии цвет оценивается данными среднестатистического наблюдателя большой группы здоровых людей)
Понятие зрительная кора включает первичную зрительную кору (также называемую стриарной корой или зрительной зоной V1) и экстрастриарную зрительную кору — зоны V2, V3, V4, и V5. (См. о зонах V2, V3, V4, и V5 в статье Зрительная кора)
Математика цветного зрения[править | править код]
Сравнение теорий цветного зрения трёхкомпонентной и нелинейной[править | править код]
Сравнение теорий цветного зрения трёхкомпонентной и нелинейной обсуждается в основном на материалах исследований человека, но и опыты на животных также служат материалом для сравнений. Все данные материала представлены в виде таблиц сравнений.
Выводы[править | править код]
Анализ данных цветовосприятия в последние десятилетия показывает, что c появлением новой техники, технологии исследований, учёным удалось доказать, что принцип трихроматизма (принцип трёхкомпонентного цветного зрения) является основой цветного зрения. На базе исследований на живых клетках при воздействии на них видимых лучей света (цвета) в цвете удалось рассмотреть колбочки S,M,L (RGB), которые участвуют в процессе дневного зрения, в процессе формирования цветного изображения в зрительных отделах головного мозга в трёхмерном пространстве в цвете, в стерео. C открытием третьего фоторецептора ганглиозного слоя сетчатки (см. ipRGC (~2007 год)), который расположен не в фокальной поверхности сетчатки, синаптически связан с колбочками, палочками, мозгом. Он выполняет важные функции при цветовосприятии в регулировании работы колбочек при восприятии спектра коротковолновых лучей (синих-фиолетовых), пропуская нормальные лучи спектра с длиной волны более 498нм, перенаправляя их в зону восприятия синих лучей колбочек-S, блокируя сильнодействующие синие лучи с высокой частой колебания — длинами волн менее 498нм. Он содержит оптический пигмент меланопсин, чувствительный к синим лучам (S) света.
Проводились исследования учёными Е. О. Загальской и В. П. Гнюбкиной из Института Биологии Моря ДВО РАН (2006 год), Владивосток. Цель этого исследования состояла в том, чтобы проанализировать ретиномоторные движения у взрослых рыб (zebrafish) и процесс созревания ретиномоторной реакции в его развитии у молодняка рыб. Показано, что ретиномоторные движения используются как адаптация зрения в условиях тёмно/светового освещения, показан механизм адаптации у рыбы-зебры. У взрослых особей гранулы меланоцитов.

В 2006[78] году при помощи трансмиссионной электронной микроскопии пигментного слоя сетчатки исследован эпителий клеток сетчатки глаза, где вырабатывается пигмент меланин (коричневого и чёрного цвета), который обеспечивает открытие и закрытие зон освещения колбочек и палочек в зависимости от вида освещения. При дневном освещении данный пигмент блокирует работу палочек, которые ко всему ещё уходят в зону под колбочки. И наоборот, при сумеречном освещении палочки выходят из под прикрытия зоны прохождения синих и ультрафиолетовых лучей меланином, а колбочки опускаются в зону ниже палочек и находятся под прикрытием меланина. (См. рис. C). В конечном итоге подтверждено открытие явления ретиномоторной реакции фоторецепторов , работы колбочек и палочек в условиях дневного и ночного освещения учёными Е. О. Загальской и В. П. Гнюбкиной из Института Биологии Моря ДВО РАН, Владивосток.
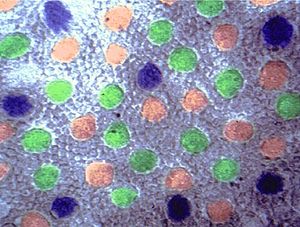 Рис. S. Распределение рецепторов в сетчатке бабуина . Синие колбочки в периферийной зоне ямки фовея с углом охвата 7-8 градусов были распределены регулярно, красные и зеленые колбочки были распределены беспорядочно всюду. Плотность распределения зелёных колбочек больше, чем красных, больше чем синих.[79]Рентгеноскопия сетчатки и отделов мозга, проведенная ещё в 1966, повторно в 2009 годах Р. Е. Марком, на атомно-молекулярном уровне подтверждает ранее сформулированые гипотезы о работе сетчатки, где заложены фоторецепторы колбочки, выделющие основные лучи спектра S,M,L («синие», «зелёные», «красные») (см. Фиг. R, рис. S), которые на рецепторном уровне воспринимают, трансдукцируют основные сигналы RGB оптического изображения (контурного, рецепторного) перед передачей их в высшие зрительные отделы (нейронные) головного мозга (см.Труды доктора Р.Е.Марка и его лаборатории). При этом следует подчеркнуть, что структукрно колбочки устроены одинаково, но в зависимости от места расположения они имеют разные морфологические размеры, например, в центральной ямке сетчатки они самые маленькие, так как плотно упакованы и при этом, в зоне ямочки диаметром до 1 мм расположены только колбочки красные и зелёные, где нет палочек вообще. Каждая колбочка способна оппонентно выделить один из основных цветовых сигналов S,M,L, RGB, так как фотопигмент опсин в зависимости от сфокусированного и выделенного основного луча предметной точи способен в результате мутации белка пигмента принимать красную, зелёную и синюю окраску!
Рис. S. Распределение рецепторов в сетчатке бабуина . Синие колбочки в периферийной зоне ямки фовея с углом охвата 7-8 градусов были распределены регулярно, красные и зеленые колбочки были распределены беспорядочно всюду. Плотность распределения зелёных колбочек больше, чем красных, больше чем синих.[79]Рентгеноскопия сетчатки и отделов мозга, проведенная ещё в 1966, повторно в 2009 годах Р. Е. Марком, на атомно-молекулярном уровне подтверждает ранее сформулированые гипотезы о работе сетчатки, где заложены фоторецепторы колбочки, выделющие основные лучи спектра S,M,L («синие», «зелёные», «красные») (см. Фиг. R, рис. S), которые на рецепторном уровне воспринимают, трансдукцируют основные сигналы RGB оптического изображения (контурного, рецепторного) перед передачей их в высшие зрительные отделы (нейронные) головного мозга (см.Труды доктора Р.Е.Марка и его лаборатории). При этом следует подчеркнуть, что структукрно колбочки устроены одинаково, но в зависимости от места расположения они имеют разные морфологические размеры, например, в центральной ямке сетчатки они самые маленькие, так как плотно упакованы и при этом, в зоне ямочки диаметром до 1 мм расположены только колбочки красные и зелёные, где нет палочек вообще. Каждая колбочка способна оппонентно выделить один из основных цветовых сигналов S,M,L, RGB, так как фотопигмент опсин в зависимости от сфокусированного и выделенного основного луча предметной точи способен в результате мутации белка пигмента принимать красную, зелёную и синюю окраску!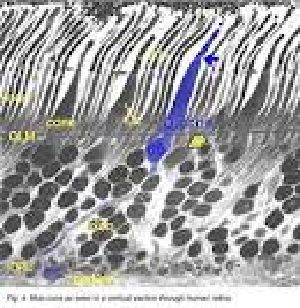 Рис. 5. Синяя колбочка-S в вертикальном разрезе через сетчатку человека. В вертикальных срезах сетчатки человека S-колбочка распознается наличием более длинного внутреннего сегмента, который больше прилипает к субретинальному пространству, чем соседние конусы (фиг.4).[80]
Рис. 5. Синяя колбочка-S в вертикальном разрезе через сетчатку человека. В вертикальных срезах сетчатки человека S-колбочка распознается наличием более длинного внутреннего сегмента, который больше прилипает к субретинальному пространству, чем соседние конусы (фиг.4).[80]


В настоящее время уже вообще не стоит вопрос о восприятии цвета в сетчатке блоками из трёх колбочек S,M,L (R.Marc, 1977).[79].[81] Если говорить о восприятии синих монолучей синими колбочками, то уже в 1977 году получены легко рентгеновские снимки в плане фокальной поверхности сетчатки синей колбочки в ряду основных колбочек зелёных и красных (см. рис. S).
Группа учёных Лаборатории R.Marcа (Лаборатория доктора Марка, Труды доктора Р.Е.Марка и его лаборатории) уже работают над созданием атласа основных нервных клеток сетчатки и отделов головного мозга, на основании электронной микроскопии их у кроликов и крыс. При этом используется разработаная мощная программа обработки данных современных микроскопов и компьютерных систем. При этом открывается возможность визуально анализировать работу здоровых и больных клеток с целью лечения целого ряда болезней (слепота, дальтонизм и др.[79]
Что важно, на рентгеноскопии среза сетчатки (cм. Фиг. R) видно, что основные фоторецепторы колбочки и палочки в процессе формирования оптического изображения работают независимо. Так синяя колбочка при дневном зрении воспринимает диапазон синих лучей под присмотром фоторецепторов ipRGC. Например, сильные сигналы ультрафиолетовых, синих лучей под действием ipRGC и мозга закрывают вообще ресницами глаза глаз от их попадания во внутрь (скорость реакции 1/2000сек) и имеются другие функции у них, связанные с трансдукцией синего сигнала.
- Доводы о том, что палочки и колбочки одинаковы и не имеют различий голословны и оторваны от жизни. В настоящее время получены изображения разных типов колбочек и палочек, которые отличаются морфологически и функционально. (См. рис. 26). На приведенных изображениях колбочек птиц и черепах ещё раз показан вид колбочек, которые морфологически одинаковы, но различаются видом, в зависимости от места расположения и меняющемуся виду при работе. В глазу птиц даже есть двуглавые колбочки, расположенные в периферийной части и управляющие движением птиц.

Что касается фотопигментов, то следует знать, что пигмент, например, фоторецептора или изменяют своё устройство или претерпевают уменьшение своего изображения (вида), когда они поглощают фотон. Свет преобразует зрительный пигмент через следующий ферментный каскад: фотоны — родопсин — активизированный rhodopsin (metarhodopsin II) — GTP обязательный белок (трансдуцин) — фермент, гидролизирующий cGMP (cGMP-phosphodiesterase) — в итоге закрывающаяся мембрана, связавшая cGMP-регулируемым катионным каналом. (См. рис.9).
В темноте устойчивый поток течет в открытые каналы, которые несут главным образом ионы Na, составляя «темный поток», который частично деполяризует ячейку фоторецептора. Таким образом, деполяризованный фоторецептор выпускает медиатор (предполагая, что это является глутаматом аминокислоты) из его синаптических терминалов на нейроны второго заказа в темноте. На легком (световом) возбуждении rhodopsin молекулы — подводят isomerized к активной форме, вышеупомянутого следующего каскада, приводя к закрытию каналов катиона мембраны фоторецептора, останавливая темный поток и заставляя мембрану клетки фоторецептора гиперполяризовать и останавливаит выпуск медиатора к нейронам второго заказа (см. Stryer, 1991; Yau, 1994, и Kawamura, 1995, для обзоров.[83]
Это изменение происходит в устройстве или в окислительно-восстановительном процессе хромофора и выражается в изменении устройства белка или его деятельности, и вызывает каскад трансдукции сигнала. Примеры для пигментов фоторецепторов, находящиеся в сетчатке глаза: например, родопсин (относящийся к сложным белкам хромопротеинам) вызывает жёлтый цвет; например, криптохром (cryptochrome), и билин (bilin) вызывает голубой цвет растений. Относительно фотопигмента цианолаба, который ещё не выделен учёными, то в результате фототрансдукции сигнала, пигмент, который светится в жировых капельках синим цветом, увиден и его можно назвать любым именем, важен цвет, который виден при исследованиях на снимках, и который имеется на синих колбочках-S. Не исключено, что он близок к меланопсину, билину. В любом случае, он является разновидностью фотопигмента опсина (См. Зрительные пигменты и передача зрительного сигнала).[84]. [цитата, необходимая]
- По вопросу хроматической аберрации, визуального восприятия предметных точек, остроты зрения, о строении фокальной поверхности сетчатки (мозаика, блоки фоторецепторов) и др.
.JPG)
Далее, например, с точки зрения нелинейной теории зрения, при исследовании по методу С.Ременко с главным принципом цветного зрения, где участвуют палочки с колбочками, не возможно получить объективные данные ещё по следующим причинам (см. рис. R,9,1,1г,8b,1p):
- 1) Не возможно дифференциально оценить работу колбочек в местах их расположения при сканирование фоторецепторов оптического изображения на фокальной поверхности сетчатки;
- 2) (См. рис. 1p). Получить и оценить сфокусированную предметную точку в области жёлтого пятна. Так как там нет палочек вокруг колбочек, что сразу исключается главный вывод нелинейной теории зрения, когда утверждается, что при цветном зрении работают палочки вместе с колбочками. А если учесть, что в жёлтом пятне около 97 % колбочек, то какая речь может идти о цветном зрении по принципу С.Ременко. Синие же лучи воспринимаются в этой центральной фовеальной ямке с 3,5 % колбочками, расположенными в поясе за пределами центра фовеальной ямки. Откуда построенные графики не соответствуют графикам, построенным при иследовании на срезах и фронтальной поверхностях живой сетчатки.
Рассуждения об аберрации основных лучей спектра света при фокусировании в трёх сечениях колбочек с выделенными тремя основными лучами RGB исключаются по нескольким причинам:
- колбочки в центральной ямке (M,L) работают только с двумя лучами S,M,L или с одним лучом S. (Синие лучи попадая в эту зону под управлением горизонтальных фоторецепторов ipRGC, при сканировании таких сфокусированных предметных точек в зоне жёлтого пятна с радиусом более 0,13 мм, с большой скоростью в фемтосекунды трансдуктируются в ближайшую колбочку-S (синюю) (их около 3,5 %), расположеннную в не зоны центра с радиусом 0,1 мм).
- вообще, колбочки работают индивидуально, но в блоках по три S,M,L, которые при фокусировании предметных точек на фокальную поверхность накрываются кружком нерезкости с диаметром 7-9 мкм в любом месте сетчатки в зону области (400-700)нм с центром 550нм, привязанной к центру центральной ямки L. Оппонентно выделяемый биосигнал сфокусировыанного луча на любую независимую колбочку блока S,M,L отпрвляется в мозг.
При этом как известно, при выделении основных зрительных сигналов S,M,L всех точек оптического изображения на сетчатке для отправки в мозг происходит сложнейший оппонентный отбор из трёх пар цветов: красный+зелёный, синий+жёлтый, белый+чёрный более яркого одного из основных RGB сигналов S,M,L. Этого не возможно получить на транзисторной электронной схеме при колориметрии.
- Вообще, данные ретиномоторной реакции фоторецепторов говорят о том, что в условиях дневного освещения работают только колбочки, обеспечивая цветное зрение.
- 5) Фоторецепторы сетчатки глаза ipRGС, открытых в 2007 году, и содержащих пигмент меланин в зрительном процессе выступают также как фильтр. Они управляют диафрагмированием зрачка глаза, что важно при изменении освещения. Например, при сумеречном освещении диаметр зрачка наибольший (порядка 6 мм), что позволяет палочкам вступать в работу. При сканировании оптического изображения фоторецепторы ipRGC управляют сигналами предметных точек, сфокусированных в зоне жёлтого пятна, где нет синих колбочек и палочек, которые со скоростью в фемтосекунды направляются в зону к синим колбочкам, передающих данный сигнал синего цвета в мозг. Подробнее смотри Труды доктора Р.Е.Марка и его лаборатории, Теорию трёхкомпонентного цветного зрения и т.д.
При учёте хроматических аберраций глаза учитывается высота мембраны колбочки в 50мкм. Любой сфокусированный монохроматический луч спектра может сфокусироваться на разной высоте мембраны, даже если все колбочки строго одинаково расположены в фокальной поверхности сетчатки. Однако и в этом случае мембраны колбочек на изображении среза сетчатки расположены не на одной высоте. Палочки расположены выше колбочек. Подробнее смотри выше раздел «Мозаика экстерорецепторов сетчатки». При этом вопрос аберраций света в глазе имеет фундаментальный характер цветного зрения. Так как при оппонентном выделении колбочками основных световых сигналов S,M,L, RGB в блоке, например, трёх колбочек, а сфокусированный луч предметной точки фокусируется в виде кружка нерезкости равной или больше кружка нерезкости (3‒5мкм), то в режиме оппонентного отбора наиболее ярких монолучей, каждая колбочка выделяет один из трёх основных лучей S,M,L, которые посылаются в мозг. При этом каждый сфокусированный луч ложится в конусной мембране в сечениях в зависимости от его длины волны с размером диаметра поперечного сечения фронта волны. Например, синий луч проходит до наименьшего размера сечения конуса конусной мембраны колбочки, то есть у вершины конуса. Или в пределах всей длины мембраны в 50мкм лучи фокусируются точно на разных высотах, что исключает погрешности хроматической аберрации и обеспечивает глазу выдавать биосигналы лучей предметных точек в мозг без искажений. Откуда оптическая система глаза не требует от преломляющей системы хрусталика и зрачка устройства по типу современных анастигматов как в фотографии. Где все лучи фокусируются в одной фокальной плоскости.
Дополнительно цветное зрение у людей и животных связано с вопросами саморегуляции организма, с наличием совокупного движения, поведения биологических систем, которые должны быть замкнутыми и одновременно должны иметь модулятор движения, связывающий организм и его внутренние системы с подвижностью, колебаниями внешней среды, из которой поступает сигналы, возбуждающие по амплитуде или частоте, требующие взаимодействия с элементами нервных узлов. Это относится также и к сетчатке с фоторецепторами. Живая ткань не может быть статичной и колбочки также подвержены ретиномоторной реакции, что сказывается на восприятие, на зрительный процесс в режиме дневного и сумеречного освещения (См. Ретиномоторная реакция фоторецепторов сетчатки глаза).
На основании флуоресцентной наноскопии срезов живой сетчатки доказано, что восприятие света и цвета основано на работе фоторецепторов сетчатки, состоящей из мозаики блоков («клеток») с разным количеством колбочек, которые воспринимают основные монохроматические лучи спектра света предметной точки индивидуально в зависимости от строения сетчатки. Например, у птиц мозаика сетчатки состоит из блоков, содержащих систему колбочек «четырехроматик» (четыре колбочки), у человека — «трихроматизм» (три колбочки). (см. рис.3а,6). То есть в каждом блоке мозаики сетчатки могут содержаться по три, четыре, пять и т. д. колбочек, воспринимающих и выделяющих оппонентно основные лучи S,M,L (RGB).
- Также на основании опубликованных данных исследований профессора департамента офтальмологии функциональной нейроанатомии сетчатки R.E.Marcа (2009), то уже в 1966 году при электронной микроскопии сетчатки получены данные «синих» колбочек приматов (см. фиг. К).[79](См.Труды доктора Р.Е.Марка и его лаборатории).
Например, из условия наличия 6 млн колбочек в жёлтом пятне (у человека), на площади 6 мм², которые воспринимают цвет, можно на базе известных данных показать, что одна колбочка не в состоянии выдать нужную информацию цветов, сфокусированной на сетчатку предметной точки. Известно, что разрешающая способность нормального глаза при чтении с расстояния 250 мм находится в пределах 0,072‒0,200 мм и в зависимости от освещённости и индивидуума примем среднестатистическую величину оценок разрешающей способности оптических приборов, среднестатистических глупп взрослых людей, проходящих тестирование (водителей транспортных средств, военнослужащих и т. д.) с показателем 0,0896мм (При остроте зрения 0,8). Основное количество фоторецепторов желтого пятна в центре сетчатки (~6млн) имеет площадь ~ 5,6‒6 мм², (то есть оптическое изображение содержит 1000000 (1мгп) разных цветовых точек), где: расстояние между одноименными точками (фоторецепторами или «пикселами») равно примерно 3‒4мкм (диаметральный наибольший размер основания конуса мембраны колбочки). Рассматриваемые визуально, например, две предметные точки в виде облачков, кружков нерезкости с просветом, равный диаметру кружка нерезкости, которые глаз чётко видит. (Более подробно см. Современный взгляд на световосприятие и цветное зрение, Острота зрения человека).


Из условия остроты зрения глаза — способности рассмотреть две точки с просветом между ними на определённом расстоянии (например, при остроте зрения человека 1,0 оно равно 72,5мкм), одна колбочка этого сделать не может. Откуда всегда рассматривается две точки с просветом минимум двумя колбочками или палочками, которые образуют блоки, например, S,M,L мозаики сетчатки (См. Цветное зрение у птиц). Как показано выше, диспергированные монолучи предметной точки с диаметром кружка нерезкости примерно 7мкм накрывают три колбочки минимум одного блока с диаметром 6,9мкм (где диаметр мембраны колбочек примерно = 3‒4мкм). Таким образом, в любом случае предметные точки участвуют в восприятии изображения блоком из трёх колбочек мозаики сетчатки. А это значит, что 6 млн колбочек жёлтого пятна объединены в 1,5‒2 млн блоков RGB, которые оппонентно трансдукцируют выходные биосигналы красного, зелёного, синего цвета под управлением головного мозга и фоторецепторов ipRGC, попадающие в зрительные отделы головного мозга через жировые капельки и далее по нервным путям. (Около 1 млн стволов). При этом часть сигналов из 1,5‒2 млн фильтруется, подавляется в блоках при трансдукции RGB под управлением головного мозга и фоторецепторов ipRGC. Принято, что все колбочки одинаковые, каждая колбочка работает в оппонентном отборе основных лучей спектра S,M,L (RGB) предметной точки, которые на неё сфокусированы. При этом наиболее ярких лучей.
Ссылкой на источник в виде:
- Единственные результаты, так ни кем и не оспоренные на сегодняшний день, были получены группой учёных W. B. Marks, W. H. Dobelle, E. F. Mak Nichol. Их исследование подробно описано в работе «Visual Pigments of Single Primate Cones» W. B. Marks, W. H. Dobelle, E. F. Mak Nichol Department of Biophysics, Johns Hopkins University, Baltimore, Maryland 21218. Science, Vol. 143, p. 1182, 1964., и что при освещении только её узкополосным излучением (вырезанным из широкого непрерывного спектра), по спектру отражения можно определить именно ту область, которую поглощает колбочка (область чувствительности колбочки). Для этого использовался широкополосный спектр оптического излучения из которого монохроматором вырезался очень узкий участок спектра. Далее этот монохромный спектр фокусировался в точку размером меньше, чем геометрический размер самой колбочки. Эти точки сканировали в сетчатке живого глаза.
Как видим, источник, где проведенное исследование в 1964 году при сканировании сетчатки не учитывалась работа мозга и фоторецептора ipRGC (открытые в 2007 году) с фотопигментом меланопсином, которые дозируют, фильтруют область сильных фотонов коротковолновых монохроматических лучей (фиолетовых-синих) не случайно не позволили в то время обнаружить колбочек-S, трансдукцирующих синие лучи света! (Очень ценное обобщение).
- По вопросу палитры цветов.

Треугольник Cb-Cg-Cr это только xy=(0,0), (0,1), (1,0), треугольник CIE xy— цветность пространства. Линия, соединяющая Cb и Cr это линия нулевой яркости на цветовом графике (alychne).
Обратите внимание, что спектральный локус проходит через rg=(0,0) на 435.8 нм, через rg=(0,1) в 546.1 нм и через rg=(1,0) при 700 нм.
- По вопросу палитры цветов согласно трихроматизма и теории многокомпонентного цветного зрения, которую воспринимает зрительная система глаза с учётом работы зрительных отделов мозга, критика представителей нелинейной теории выглядит:
- … Палитра цветов, которую смог бы различать глаз работающий в соответствии с трёхкомпонентной «гипотезой» зрения будет слишком мала, что совершенно не соответствует действительности. Нанесём на цветовой треугольник точки максимумов чувствительности пигментов глаза (по предположениям трёхкомпонентной «гипотезы», максимумы чувствительности предполагаемых колбочек). На рис. 4 видим область полученной цветовой палитры (внутри треугольника с углами в точках 430 нм, 540 нм и 570 нм), которую можно анализировать или синтезировать при известных максимумах чувствительности пигментов (рис. 2.).
- Согласно Рис.2a в Диаграмме CIE rg цветность пространства показывает построенный треугольник, определяющий цветовое пространство CIE XYZ.
Треугольник Cb-Cg-Cr это только xy=(0,0), (0,1), (1,0), треугольник CIE xy— цветность пространства. Линия, соединяющая Cb и Cr это линия нулевой яркости на цветовом графике (alychne). Обратите внимание, что спектральный локус проходит через rg=(0,0) на 435.8 нм, через rg=(0,1) в 546.1 нм и через rg=(1,0) при 700 нм! (По старой системе применялось цветовое пространство RGB (цветовая модель) и рассчитывалось без учёта линейной функции, а область полученной цветовой палитры была внутри треугольника с углами в точках 430 нм, 540 нм и 570 нм). Значение же энергии точки (E) при rg=xy=(1/3,1/3) равное. (См. также XYZ (цветовая модель), Цветовые координаты). Таким образом палитра цветов, которую различает глаз, работающий в соответствии с теорией многокомпонентного цветного зрения охватывает весь диапазон цветов с длинами волн rg=(0,0) на 435.8 нм, rg=(0,1) в 546.1 нм и rg=(1,0) при 700 нм! (См. рис.2a,XYZ (цветовая модель), Цветовые координаты).
- По вопросу дефектов цветовосприятия.
Именно, с открытием третьего фоторецептора сетчатки ipRGC (2007) учёные получили возможность глубже вникнуть в природу отклонений в восприятии цвета зрительной системой. Не рассматривая в комплексе работу колбочек, мозга, фоторецепторов ipRGC, конечно, не возможно оценить цветное зрение! Только с позиций трихроматизма — теории многокомпонентного цветного зрения практически решаются все вопросы цветовосприятия.
Кажется всё понятно[править | править код]
- Так сколько же типов колбочек существует в глазу человека?. Что можно сказать по вопросу критики трёхкомпонентной теории цветного зрения?
Так вот: При рентгеноскопии, микроскопии срезов живой сетчатки мы сейчач видим три независимые колбочки RGB,S,M,L (синюю, зелёную, красную) и палочку, не связанную с колбочками. Уже с 1966г., а в 2009г. повторно доказало — при цветном зрении работают только три колбочки и присутствие синей колбочки-S говорит о наличие синего пигмента цианолаба - разновидности опсина, а полочки работают при сумеречном и ночном освещении (после заката солнца) (см. Труды доктора Р.Е.Марка и его лаборатории др.).
- Что же получаем в итоге?[17]
- В итоге получаем:
- Нелинейная теория зрения С. Ременко 1975 года, основной вывод которой является, что при цветном зрении работают фоторецепторы колбочка+палочка не подтверждён ни одним методом исследований с применением рентгеноскопии, микроскопии живых клеток сетчатки, а также методами трансмиссионной электронной микроскопии пигментного слоя сетчатки, денситометрии при измерении цвета фотопигментов колбочек и т.д. Получаем, что нелинейная теория и выводы не верны. (Откуда в мире нелинейная двухкомпонентная теорию зрения С. Ременко не получила своего признания).
См. также[править | править код]
- Труды доктора Р.Е.Марка и его лаборатории
- Цветное зрение
- Визуальное цветное зрение
- Специфика работы мембраны колбочек сетчатки глаза
- Цветное зрение у человека
- Оптический тракт (версия Миг)
- Наружное коленчатое тело
- Зрение человека (версия Миг)
- Измерение цвета (версия Миг)
- Опсины (версия Миг)
- Зрительная фототрансдукция (версия Миг)
- Зрительная кора
- Острота зрения
- Теория оппонентного цветного зрения (версия Миг)
- Ретиномоторная реакция фоторецепторов сетчатки глаза
- Ночное зрение
- Дневное зрение
- Зелёный луч
- Нелинейная теория зрения
Примечания[править | править код]
- ↑ а б в Robert E. Marc. FNAR/ "FNAR" (PDF). Retrieved Лаборатория доктора Марка,2009. Check date values in:
|accessdate=(help) - ↑ а б в г John A. Medeiros. "Color Vision: A New Understanding". Cones and Color.net. Retrieved 8 сентября, 2012. Check date values in:
|accessdate=(help) - ↑ а б в г д Goldsmith, Timothy H (Июль 2006). "«What birds see» (PDF)" (PDF). Scientific American. pp. 69–75. Retrieved 8 сентября, 2012. Check date values in:
|date=, |accessdate=(help) - ↑ а б в г д е Gerald K. Huth. "THE RETINA AS AN ARRAY OF NANO-ANTENNAS". Retrieved on January 13, 2011. Check date values in:
|accessdate=(help) - ↑ http://webvision.med.utah.edu/2013/02/photonic-crystal-light-collectors-in-fish-retina-improve-vision-in-turbid-water/
- ↑ http://neurobiology.ru/res/ResourceFile/45/FILE_FILENAME/2006-10-44.pdf
- ↑ https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1438839/pdf/jrsocmed00261-0028.pdf
- ↑ http://webvision.med.utah.edu/book/part-ii-anatomy-and-physiology-of-the-retina/photoreceptors/
- ↑ Marks, W.B., Dobelle, W.H. & MacNichol, E.F. Visual pigments of single primate cones. Science (1964) 143: 1181—1183.
- ↑ https://translate.google.ru/translate?hl=ru&sl=en&u=https://www.ncbi.nlm.nih.gov/books/NBK235548/&prev=search
- ↑ Wald G., The photochemistry of vision, Doc. Ophthalmol., 3, 94 (1949)
- ↑ Günther Wyszecki Color Science: Concepts and Methods, Quantitative Data and Formulae. — 2nd. — New York: Wiley Series in Pure and Applied Optics, 1982. — ISBN 0-471-02106-7о книге
- ↑ The Reproduction of Colour. — 6th. — Chichester UK: Wiley–IS&T Series in Imaging Science and Technology, 2004. — С. 11–12. — ISBN 0-470-02425-9о книге
- ↑ Тонквист Г. Аспекты цвета. Что они значат и как могут быть использованы // Проблема цвета в психологии / Отв. ред. А. А. Митькин, Н. Н. Корж. М.: Наука, 1993. С. 5‒53. (С. 7)
- ↑ ГР Jacobs, Neitz М., Deegan JF, и Neitz J. Trichromatic окрашивают видение в Новых Мировых обезьянах. Nature. 382: 156†"158, 1996}}
- ↑ Neitz М. и Neitz J. Молекулярная генетика цветного видения и цветных дефектов видения. Арч Офталмол 118: 691†"700, 2000. [FreeВ FullВ Текст]
- ↑ http://physiologyonline.physiology.org/content/17/3/93.full
- ↑ S-Potentials and Horizontal Cells. — Webvision. — С. 688. — ISBN 5-02-000747-1о книге<ul><li>Регулярное выражение «ISBN» классифицировало значение «5020007471↑http://prometheus.med.utah.edu/~marclab/Marc_Duanes_FNAR_20080815_layout.pdf↑Goldsmith» как недопустимое.</li> <!--br--><li>Регулярное выражение «ISBN» классифицировало значение «TimothyH.(July2006).«Whatbirdssee»(PDF).ScientificAmerican:6975.http://www.csulb.edu/labs/bcl/elab/avian%20vision_intro.pdf↑http://webvision.med.utah.edu/HC.html» как недопустимое.</li></ul>
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ а б в г Michael Kalloniatis and Charles Luu. "Color Perception by Michael Kalloniatis and Charles Luu". Retrieved August 2012.
- ↑ http://thelib.ru/books/bse/bolshaya_sovetskaya_enciklopediya_cv-read-10.html
- ↑ http://en.wikipedia.org/wiki/Cone_cell
- ↑ http://en.wikipedia.org/wiki/Cone_cell
- ↑ Schacter,Gilbert, Wegner, «Psychology», New York: Worth Publishers,2009.
- ↑ Color Vision: A New Understanding.
- ↑ Finn, J. T., Grunwald, M. E, and Yau, K-W. 1996. Cyclic nucleotide-gated ion channels: An ex-tended family with diverse functions. Annu. Rev. Physiol.58: 395—426.
- ↑ Nakanishi, S., Nakajima, Y., Masu, M., Ueda, Y, Nakahara, K., Watanabe, D., Yamaguchi, S., Kawabata, S., and Okada, M. 1998. Glutamate receptors: Brain function and signal trans-duction.
- ↑ http://webvision.med.utah.edu/book/part-ii-anatomy-and-physiology-of-the-retina/photoreceptors/
- ↑ http://webvision.med.utah.edu/2013/02/photonic-crystal-light-collectors-in-fish-retina-improve-vision-in-turbid-water/
- ↑ Остерберг Г. Топография слоя стержней и конусов в сетчатке человека. Acta Ophthal Suppl. 1935; 6: 1-103.
- ↑ Curcio CA, Sloan KR, Packer O, Hendrickson AE, Kalina RE. Распределение конусов в сетчатке человека и обезьян: индивидуальная изменчивость и радиальная асимметрия. Науки. 1987; 236: 579-582. [ PubMed ]
- ↑ http://webvision.med.utah.edu/book/part-ii-anatomy-and-physiology-of-the-retina/ph
- ↑ http://prometheus.med.utah.edu/~marclab/lex_refs/marc_sperling_science_1977.pdf
- ↑ https://en.wikipedia.org/wiki/Peripheral_vision
- ↑ /translate.google.ru/translate?hl=ru&sl=en&u=https://www.ncbi.nlm.nih.gov/pubmed/11322974&
- ↑ http://prometheus.med.utah.edu/~marclab/Marc_Duanes_FNAR_20080815_layout.pdf
- ↑ (Helga Kolb) http://webvision.med.utah.edu/book/part-iii-retinal-circuits/s-cone-pathways/
- ↑ http://www.ghuth.com/2011/01/16/found-the-first-elusive-blue-cone/
- ↑ http://www.ghuth.com/
- ↑ http://webvision.med.utah.edu/book/part-iii-retinal-circuits/s-cone-pathways/
- ↑ а б Vitamin A
- ↑ GPCR
- ↑ http://commons.wikimedia.org/wiki/User:%D0%AE%D0%BA%D0%B0%D1%82%D0%B0%D0%BD#mediaviewer/File:Retina_layers.svg
- ↑ http://prometheus.med.utah.edu/~marclab/Marc_Duanes_FNAR_20080815_layout.pdf
- ↑ Рецептор
- ↑ Фоторецептор
- ↑ Wald G., The photochemistry of vision, Doc. Ophthalmol., 3, 94 (1949)
- ↑ http://physiologyonline.physiology.org/content/17/3/93
- ↑ http://ru.wikipedia.org.advanc.io/wiki/%D0%90%D1%83%D1%82%D0%BE%D1%81%D0%BE%D0%BC%D0%B0
- ↑ http://en.wikipedia.org/wiki/File:Fig_retine.png
- ↑ Wong, Kwoon Y.; Dunn, Felice A.; Berson, David M. (22 декабря, 2005). "«Photoreceptor Adaptation in Intrinsically Photosensitive Retinal Ganglion Cells» (HTML: Full text)" 48. Neuron. pp. 1001—1010. doi:10.1016/j.neuron.2005.11.016. Retrieved 11 мая, 2011. Check date values in:
|date=, |accessdate=(help) - ↑ http://en.wikipedia.org/wiki/Photopigment
- ↑ http://en.wikipedia.org/wiki/Melanopsin
- ↑ http://en.wikipedia.org/wiki/Retina
- ↑ http://en.wikipedia.org/wiki/Ganglion_cell
- ↑ http://en.wikipedia.org/wiki/Photosensitive Фотосенсибилизация
- ↑ http://www.conesandcolor.net/_E_Cone_Shape.htm
- ↑ http://www.conesandcolor.net/_E_Cone_Shape.htm
- ↑ http://www.imls.uzh.ch/static/CMS_publications/neuhauss/literatur/pdf06/Hodel_AnatRec_06.pdf
- ↑ http://www.imls.uzh.ch/static/CMS_publications/neuhauss/literatur/pdf06/Hodel_AnatRec_06.pdf
- ↑ Pierce ME, Besharse JC (1985) Circadian regulation of retinomotor movements. I. Interaction of melatonin and dopamine in the control of cone length. J Gen Physiol 86:671‒689
- ↑ http://www.molvis.org/molvis/v14/a44 Identification of differentially expressed genes in carp rods and cones Molecular Vision 2008; 14:358‒369
- ↑ http://en.wikipedia.org/wiki/Color_vision
- ↑ Натанс, Джереми; Томас, Дарси Hogness, Давид Сергеевич (11 Апреля, 1986). «Молекулярная Генетика Человека Цветового Зрения: Гены, кодирующие Синие, Зеленые и Красные, Пигменты». Наука 232 (4747): 193—202. Bibcode:1986Sci…232 193N… doi:10.1126/science.2937147. JSTOR 169687. PMID 2937147.
- ↑ Нейтц Дж., Якобс GH (1986). «Полиморфизм длинноволнового конуса в нормальное цветовое зрение человека». Природа 323 (6089): 623-5. Bibcode:1986Natur.323 623N… doi:10.1038/323623a0. PMID 3773989.
- ↑ Jacobs GH (январь 1996 года). «Предстоятель photopigments приматов и цветового зрения». Proc. Natl. Acad. Sci. США 93 (2): 577-81. Bibcode:1996PNAS…577J 93… doi:10.1073/pnas.93.2.577. PMC 40094. PMID 8570598.
- ↑ http://en.wikipedia.org/wiki/Color_vision
- ↑ Трихроматия
- ↑ http://physiologyonline.physiology.org/content/17/3/93.full.
- ↑ http://webvision.med.utah.edu/book/part-ii-anatomy-and-physiology-of-the-retina/photoreceptors/
- ↑ Робинсон, С. Р. (1994). «В начале цветового зрения позвоночных». Природа 367: 121. doi:10.1038/367121a0
- ↑ http://en.wikipedia.org.advanc.io/wiki/Oil_droplet
- ↑ Часть III. Общая и специальная сенсорная физиология // Физиология человека: в 3-х томах = Human Physiology. Ed. by R.F. Schmidt, G. Thews. 2nd, completely revised edition (translated from German by M.A. Biederman-Thorson) . Т. 1. Пер. с англ.. — изд-е 2-е, перераб. и дополн.. — М.: Мир, 1996. — С. 178-321. — 323 с. — ISBN 5-03-002545-6о книге
- ↑ http://biology.ru/textbook/chapter10/section3/paragraph7/
- ↑ http://eps.dvo.ru/bm/2006/1/pdf/bm-055-059.pdf
- ↑ а б в г "Seeing in color". Prometheus. Retrieved 8 сентября, 2012. Check date values in:
|accessdate=(help) - ↑ http://webvision.med.utah.edu/book/part-iii-retinal-circuits/s-cone-pathways/
- ↑ https://www.ncbi.nlm.nih.gov/pubmed/403607?dopt=Abstract
- ↑ "Visual pigments and visual transduction.". Photoreceptors by Helga Kolb. Retrieved 17 февраля, 2014. Check date values in:
|accessdate=(help) - ↑ http://webvision.med.utah.edu/photo1.html
- ↑ http://en.wikipedia.org/wiki/Photopigment
- ↑ http://www.ghuth.com/2011/01/16/found-the-first-elusive-blue-cone/





{kind=link}
{kind=link}
{kind=link}
| Цвета и оттенки | |||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 |
|  | |||||||||||||||||||||||||||||||||||
| ∘ ∘ ∘ | |||||||||||||||||||||||||||||||||||||