Нелинейная теория зрения
Нелинейная теория зрения или точнее Нелинейная двухкомпонентная теория цветовосприятия - теория созданная С. Ременко в 1975 году, позволяющая реально описать и смоделировать процессы зрительного восприятия, ликвидировав недостатки наиболее распространённой в настоящее время трёхкомпонентной гипотезы цветового зрения.
Нелинейная модель позволяет дать четкие определения колориметрических характеристик излучения, основой которых являются физические закономерности, выражаемые математическим языком.
На основе принципов сформулированных нелинейной теорией цветового зрения построена простая действующая фотоэлектрическая модель глаза, способная однозначно распознавать любые цвета и оттенки.
Нелинейная модель цветовосприятия созданная С. Ременко, просто и логично объясняет все свойства нашего зрения не ссылаясь на участие мозга, как этого требуют другие теории и гипотезы зрения.
Введение[править | править код]
Существующая на сегодняшний день трёхкомпонентная гипотеза цветовосприятия часто преподносимая как «основная» или «общепринятая», крайне не совершенна. Она не может описать ни сам процесс цветовосприятия, ни цветовую адаптацию (баланс белого), ни нарушения в цветовосприятии при цветоаномалиях, ни множество различных эффектов, явлений и свойств нашего зрения. Все эти «пробелы» сторонники трёхкомпонентной гипотезы относят исключительно к работе головного мозга.
Конец ХХ века ознаменовался сменой научной парадигмы в области сенсорного восприятия. Было установлено что простейшие, линейные модели, а также модели, основанные на предположении о существовании определённого числа элементарных сенсоров/ощущений (в каждом из типов ощущений) не могут даже приблизительно описать принципы работы рецепторов. Например, крахом увенчались попытки свести все вкусовые ощущения — к наличию лишь 4-х рецепторов вкуса; или обонятельные — к семи типам запахов, и т.д.
Многие видные учёные, занимающиеся проблемой цветовосприятия отмечали несостоятельность трёхкомпонентной гипотезы цветового зрения. Вот только несколько примеров.
В одной из своих работ, лауреат Нобелевской премии Джордж Уолд (Нобелевская премия по физиологии и медицине, 1967 г.), в книге выпущенной журналом «Scientific American» «Восприятие, механизмы и модели» (перевод с английского Л. Я. Белополянского и Ю. И. Лашкевича. Под редакцией Н. Ю. Алексеенко. Москва, издательство «Мир», 1974 год.) на странице 138 (второй абзац снизу) признаёт: «…Споры возникают на почве незнания и мы много лет спорили о том, сколько видов колбочек участвуют в цветовом зрении человека. Многие исследователи предпочитают три, другие четыре, а один из моих английских коллег — семь. Сам я склоняюсь к трём. Это хорошее число и на сегодняшний день достаточное…». [1] Видно, что лауреат Нобелевской премии по физиологии и медицине, Дж. Уолд не может утверждать, что колбочек — именно три. Он просто склонен так думать…
Ещё одно мнение о трёхкомпонентной теории зрения — лауреата Нобелевской премии по физике 1965 года Ричарда Филлипса Фейнмана. В своих знаменитых «Фейнмановских Лекциях по Физике» в томе № 3, гл. 35 (Цветовое зрение), на стр. 175 он пишет: «Уже первые теории зрения утверждали, что имеются три сорта пигментов и соответственно три сорта колбочек, каждая из которых содержит один пигмент; что от каждой колбочки в мозг идут нервы, так что в мозг переносятся три сорта информации и там что-то происходит. Конечно, это очень несовершенная теория, ибо она не позволяет обнаружить, что за информация переносится в мозг; она даже не приступила к решению этой проблемы…». [2]
Необходимо упомянуть и мнение общепризнанного специалиста в области цветовосприятия Дэвида Хантер Хьюбела (David Hunter Hubel) — канадского и американского нейрофизиолога, лауреата Нобелевской премии по физиологии и медицине за 1981 год «За открытия, касающиеся принципов переработки информации в нейронных структурах». В своей книге (которая до сих пор служит учебным пособием по физиологии зрения и зрительного восприятия) "Глаз, мозг, зрение" он обобщает современные представления о том, как устроены нейронные структуры зрительной системы, включая кору головного мозга, и как они перерабатывают зрительную информацию. При этом в главе Цвет и зрение (в конце раздела "Теория Геринга") он пишет о своём отношении к противоречиям трёхкомпонентных и оппонентных теорий: «Что нас сегодня изумляет, так это то, что Геринг, опираясь на столь ограниченный фактический материал, сумел сформулировать теорию, так хорошо согласующуюся с нейронной организацией центральных механизмов цветового зрения. Тем не менее специалисты по цветовому зрению все еще разделены на два лагеря: одни считают Геринга пророком, а другие видят в упомянутом соответствии лишь случайную удачу. Я наверняка наживу врагов среди тех и других, так как занимаю нейтральную позицию и только чуть-чуть склоняюсь в пользу первого мнения».[3] Дэвид Хьюбел прямо признаёт, что только чуть-чуть склоняется в пользу трёхкомпонентной теории не отвергая при этом другие теории.
Нелинейная двухэлементная модель цветовосприятия[править | править код]
В 1975 году советским учёным С. Ременко была сформулирована и представлена нелинейная теория зрения[4], в основе которой заложены два принципа:
- Имеются только два типа светочувствительных элементов сетчатки глаза — однотипные колбочки и палочки;
- Осуществляется нелинейность процессов формирования сигналов цветности (без суммирования или вычитания).
Основные положения, составляющие условие задачи, были жёсткими. Допущения, которые легли в основу трёхкомпонентной теории, были исключены, и главное условие сводилось к следующему: теория должна строиться только на объективных, достоверно установленных экспериментальных данных, основываться на конкретных физических законах, без каких-либо приближений. Модель должна быть объективной и описываться математическими зависимостями в реальном трёхмерном пространстве. Электронная модель, построенная на основе принципов заложенных в теории, должна моделировать все известные явления, иллюзии и «парадоксы» цветового зрения.
Предпосылки для формулирования основных положений теории[править | править код]
Белый свет состоит из смеси электромагнитных колебаний различных длин волн. Некоторые из них поглощаются предметом, некоторые отражаются, в результате чего возникает ощущение цвета. Если пропущенный через призму свет ограничить узкой щелью и выделить поочередно цвета, то каждому цвету будет соответствовать определенная длина волны. Такой цвет будет монохроматическим - «одноцветным», Смесь двух монохроматических цветов дает ощущение другого цвета. Этот цвет может содержаться в радуге, а может и не содержаться (например, пурпурный). Жёлтый цвет может быть как монохроматическим, полученным с помощью призмы, так и получен смесью двух монохроматических цветов зелёного и красного.
Если полученный призмой жёлтый луч попытаться ещё раз разложить с помощью второй призмы, ничего не изменится. Если же через вторую призму пропустить составленный жёлтый цвет, он разложится на первоначальные красный и зелёный. Согласно законам физики два электромагнитных колебания при переходе в другую среду могут смешиваться. При этом результирующих излучений будет два: для первого частота равна - сумме частот исходных излучений, для второго - разности частот. Следовательно в случае смешения красного и зелёного, согласно длинам волн, первое излучение будет - ультрафиолетовое, невидимое; второе - инфракрасное, также невидимое. Казалось бы, результирующая смесь должна быть чёрной, а она - жёлтая! Из этого следует, что глаз воспринимает не смесь излучений, а их одновременное воздействие. Причём, если изменять интенсивность составляющих (красного и зелёного), цвет будет меняться: его можно сделать оранжевым, можно - жёлто-зелёным. При этом длины волн исходных излучений не меняются!
Таким образом, хотя в случае монохроматического излучения каждой волне соответствует определенный цвет, физически цвет ничего общего с длиной волны не имеет.
На этом основании можно сформулировать положение: в математических соотношениях, описывающих механизм восприятия излучений и преобразования результата в сигнал, представляющий собой информацию о цвете, физическая размерность частоты должна сокращаться. Следовательно цвет - величина безразмерная.
Следует принять, основываясь на предыдущих выводах: законы смешения красок и создание таким способом нового цвета - СИНТЕЗА цвета - в корне отличаются от законов восприятия глазом и опознавания мозгом цвета - АНАЛИЗА цвета. При решении проблемы цвета переносить колориметрические принципы производства красителей или фильтров на принцип биологического объекта (глаза) - неверно.
Основные положения теории.[править | править код]
Первое: цвет должен выражаться конкретным, объективным физическим параметром. Законы физики едины как для неживой, так и для живой материи. Мозг в переработке сигнала цветности участия не принимает.
Второе: так, как воспринимаются и цвета отсутствующие в спектре, длина волны не может служить параметром для определения цвета, а её размерность в формуле, определяющей сигнал цветности, сокращается. Исходя из этого, следует, что в данной формуле размерность длины волны должна находиться и в числителе, и в знаменателе. Таким образом, сигнал цветности представляет функцию от отношения сигналов двух, четырёх или любого чётного числа фоторецепторов. Вычисление сигнала цветности должно производиться нелинейными соотношениями.

Был проделан исключительно тонкий эксперимент: электроды поочередно вводились в разные точки одной и той же клетки так, чтоб клетка оставалась живой. Результат оказался следующим: в одном участке клетка реагировала на появление света (фронт импульса), в другом — на его пропадание (спад импульса). Таким образом, было установлено, что нервная клетка имеет «он» область (центр включения) и «офф» область (центр выключения). Эти области оказались разделенными участком, где вообще нет никакой реакции — ни на включение, ни на выключение. Потенциал на нём нулевой, но по одну его сторону положительный, по другую — отрицательный. Вблизи «нулевой» области сигналы малы, а по мере удаления от нее возрастают, достигают максимума, затем спадают к краям клетки. Графически это изображено на рис. 1. Кривая на этом графике изображает нелинейную функцию потенциала от расстояния, на котором находился электрод, от границы между «он» и «офф» — областями, т. е. отклонения от нулевой точки.
В технике такой механизм начал использоваться более полустолетия назад, а в настоящее время он есть в каждом доме. Это «детектор отношений», имеющийся в каждом ультракоротковолновом приёмнике, и в канале звукового сопровождения телевизора. Работает он не в световом, а в радиочастотном диапазоне электромагнитных волн. Представляет собой систему из двух контурных катушек, одна из которых настроена на длину волны короче несущей (основной длины волны радиостанции), другая — на длину волны длиннее несущей. Настройка каждой произведена так, что потенциал на них при настройке на основную длину волны одинаков и на резисторе нагрузки он равен нулю. Если длина волны передатчика изменится, то потенциал на одном из контуров увеличится, на другом - уменьшится и на нагрузке появится напряжение, зависящее от отношения этих сигналов и определяющее, насколько длина волны отклонилась от начальной, несущей.
Заметим интересную и важную особенность: так как выходной сигнал представляет функцию от отношения потенциалов, то, если мощность сигнала, приходящего от передатчика, увеличится, возрастает потенциал на контурах, причем на каждом в одинаковой степени. Вместе с тем напряжение на нагрузке не изменится, так как во сколько раз увеличится числитель, во столько увеличится и знаменатель, а одинаковые сомножители в дроби сокращаются. Здесь можно вспомнить одно из свойств зрения: изменение интенсивности освещения практически не влияет на ощущаемый цвет.

Обратим внимание ещё на одну особенность детектора отношений. Один из его контуров настроен на длину волны чуть длиннее, другой - чуть короче, напряжение на нагрузке определяет значительно большее отклонение длины волны передатчика по сравнению с длиной волны настройки контуров вплоть до длин волн, при которых на контурах образуется хоть какой-то потенциал. При этом можно заметить: чем хуже качество контуров, тем больший диапазон длин волн воспринимает система. Для радиоприёмника это не очень хорошо. Если полоса пропускания шире, чем необходимо, будут «пролезать» помехи от других станций. Поэтому полоса пропускания выбирается достаточной для передачи информации (для моно передач — уже, для стерео вещания — шире), но так, чтобы влияние помех вне рабочего диапазона было минимальным. Для этого используются фильтры, заграждающие прохождение длин волн не используемых в передаче, ограничивающие полосу пропускания.
Итак, морфологические и электрофизиологические данные говорят не только о возможности, но и о единственно возможном механизме типа «детектора отношений». Далее следует выяснить возможности этого механизма в формировании сигнала цветности.
В радиотехническом устройстве вовсе нет необходимости использовать два отдельных контура. Достаточно одного, составленного из двух половин. Для видимого света также не обязательно, скорее даже нежелательно иметь два светоприёмника, по двум причинам. При размерах в доли миллиметра невероятно трудно расположить должным образом эти приёмники. Если же они будут достаточно большими, разрешающая способность, четкость, окажется плохой. В случае же электрохимических фотоприёмников взаимовлияние двух рядом расположенных, тем более разных по составу, окажется очень сильным. Помехи, наведённые возбуждённым приёмником на невозбуждённый, будут вызывать появление противофазного, обратного сигнала, который будет ослаблять чувствительность первого. Эти помехи окажутся значительными.

Отмечен крупный «недостаток» хрусталика глаза - хроматическая аберрация. Если принять во внимание аберрацию, окажется, что оптимальным будет использование не двух отдельных, а одного, разделенного на две части фотоприёмника. Если такой приёмник расположен за призмой или линзой, то, оказывается, что нет необходимости даже специально настраивать его на соответствующие длины волн. Достаточно его размеры, место контакта и расположение выбрать соответствующим образом. Это хорошо видно на рис. 2, а. В результате преломления «красные» лучи будут падать на красночувствительную область, «зелёные» - на зелёночувствительную, «жёлтые» — в одинаковой степени на обе.
Из рисунка (рис. 2) видно, что ток, проходящий через отвод Жёл, сделанный против места падения жёлтых лучей, будет определять, на какую область падает больше света и во сколько раз. Если на каждую область падает одинаковое количество света, ток будет отсутствовать. Ток, проходящий «насквозь» через выводы Кр и Зел, определит суммарное количество света, падающего на приёмник. При условиях, показанных на рисунке, максимум соответствует жёлтой области спектра. Приёмник такой конструкции одновременно сообщает, сколько на него попадает красного, зелёного и жёлтого света и в каком соотношении (соотношение красного и зелёного). В принципе такой фоторецептор не нуждается в светофильтрах или пигментах (красителях), настраивающих его на соответствующие области спектра, хотя наличие красителей желательно для увеличения чувствительности.
Если светоприёмник расположить вдоль оси направления света, то раскладывать излучение в спектр нет необходимости. Для этого достаточно использовать низкокачественную линзу с большой хроматической аберрацией, например хрусталик. Его «недостаток» окажется не только полезным, но и необходимым. Это хорошо видно на рис. 2, b.
Ближе всего к объективу — хрусталику, располагается зелёночувствительная область, центр которой расположен в фокусе зеленых лучей. Красночувствительная область находится дальше. При этом используется ещё одно физическое свойство веществ: более коротковолновые лучи поглощаются быстрее; длинноволновые же, для того чтобы их поглотилось столько же, должны пробежать путь длиннее. В случае продольного расположения приёмника желательно наличие пигментов, усиливающих поглощение. Во-первых, белки, из которых состоит светорецептор, прозрачны в видимой области. Во-вторых, пигменты могут использоваться для автоматического регулирования чувствительности. Этот процесс называется адаптацией и рассматривается далее. Благодаря пигментам зелёночувствительная область одновременно становится более чувствительной к зелёной области спектра (пигмент хлоролаб) и является фильтром, задерживающим зелёные лучи и пропускающим красные в красночувствительную область, в которой размещён пигмент, поглощающий красные лучи (пигмент эритролаб).
Такой приёмник может считаться оптимальным и может давать достаточное количество информации как о количественном, так и о спектральном составе света.
Имеется ли похожее по конструкции устройство в сетчатке глаза?
Если вспомнить описание строения сетчатки, то такое устройство очень напоминает колбочка. Как установил Маркс с сотрудниками[5], зелёно- и красночувствительный пигменты могут сосуществовать в одной и той же колбочке. Если бы колбочка не была дифференциальным элементом, сосуществование пигментов сводило бы на нет её смысл. Колбочка отличается от палочки наличием дендритных отводов, что тоже говорит в пользу дифференциальной модели. Что, если бы в сетчатке были только такие приёмники? Как видел бы такой глаз цвет? Оказывается, свойства такой модели совпадают со свойствами глаза, в котором только Колбочки. Это явление (цветоаномалия 3-го рода, тританопия), свойственное некоторым людям, описано далее.
Итак, целесообразно ли наличие трёх типов колбочек, когда достаточно одного? Нелинейную гипотезу цветового зрения устраивает наличие только одного типа колбочек и, конечно, палочек.
Для описания моделирования эффектов нормального цветового зрения требуется, как отмечал Эвальд Геринг, автор оппонентной теории цветового зрения, ещё одна «противоположная» пара: жёлтый — синий. Необходима ещё одна колбочка? Целесообразно ли это?
Мы уже говорили, что рядом расположенные подобные элементы неизбежно будут создавать друг другу помехи. Да и нужен ли ещё один дифференциальный приёмник? Сигнал, максимум которого находится в жёлтой области, уже имеется. Это суммарный сигнал дифференциального приёмника — колбочки. Если рядом с колбочкой поместить простейший, синечувствительный приёмник, то его сигнал вместе с «жёлтым» сигналом колбочки создаст вторую пару: жёлтый — синий.
Может ли простой синечувствительный рецептор самостоятельно, без колбочки выдать цветовой сигнал? Конечно, нет. Действительно, глаз, в котором только палочки, цвета не ощущает. Может ли палочка играть роль синечувствительного элемента? Во-первых, она длиннее колбочки и выдвинута вперед. Во-вторых, она не имеет дендритных отводов. В-третьих, она содержит пигмент родопсин, увеличивающий поглощение в ней синей и зелёной областей, притом если освещение слабое — в жёлто-зелёной, если достаточно сильное — в синей. Правда, чувствительность в ультрафиолетовой области тоже высока, но эти лучи задерживаются роговицей и хрусталиком (об изменении области чувствительности более подробно будет рассмотрено далее. Здесь рассматриваются только условия достаточных освещённостей).

Для выделения дифференциального сигнала жёлтый — синий необходим смеситель. Роль его выполняют нейроны, к которым «подключены» и колбочки, и палочки. Функциональная схема такой элементарной ячейки показана на рис. 3. Этой схеме для полной ясности не хватает одного. Противоположная пара жёлтый — синий в смеси дает белый свет, а пары зелёный — красный нет. Синий, зелёный и жёлтый — спектральные тона. Если бы четвертый (противоположный зелёному) цвет был бы также спектральным, система была бы очень красивой. Однако чистый красный противоположным быть не может. И вот почему.

И рецепторы, и нейроны, и нервные волокна состоят из белков. И те, и другие поглощают интенсивно ультрафиолетовые лучи и немного сине-фиолетовые. Не удивительно, что была обнаружена реакция нейронов на свет. Часть попавшего в глаз синего света поглотится нейронами. «Синий» сигнал не изменится, «зелёный» пренебрежительно мал, а вот «красный» изменится заметно. К «красному» сигналу окажется подмешанным и «синий» сигнал нейрона.
Таким образом, при воздействии на колбочку белого света противоположным зелёному окажется не красный, а красно-синий, пурпурный. Таким путем в цветоощущение человека вошёл пурпурный цвет, которого в спектре нет. Это свойство колбочек видно из приведенной на рис. 3а кривой поглощения, снятой для «красной» чувствительности колбочки. Если бы на кривой отсутствовал «синий хвост» красночувствительной области, противоположным, дополнительным и контрастным зелёному был бы не пурпурный, а красный цвет. С учетом «синего хвоста» система приобретает изящный вид.
Итак, элементарная система из одной колбочки и одной палочки при условии, что процессы анализа цвета являются нелинейными, обладает всеми биофизическими свойствами, необходимыми для переработки световой информации и формирования сигналов яркости и цветности, которые без дополнительной переработки достаточны для однозначного опознавания цвета мозгом.
Вообще все элементы сетчатки каким-либо образом связаны между собой, и по этим связям производится обмен информацией и энергией. Обмен энергией по этим связям устанавливается не мгновенно. Такого рода зависимости от времени называются переходными процессами. Так как большинство явлений можно рассматривать только с учетом связей между элементами, такие явления, как «последовательный образ» и другие, рассматриваются совместно с переходными процессами.
Модель цветовосприятия, построенную на нелинейной гипотезе, можно представить в упрощенном виде так.
Свет, отражённый от объекта или прошедший через него, попадает на роговицу глаза и хрусталик, которые отфильтровывают губительно действующие на сетчатку ультрафиолетовые лучи. Далее свет проходит через стекловидное тело, которое задерживает содержащие много помех и мало информации инфракрасные лучи. Оставшиеся лучи, разложенные в спектр вследствие преломления роговицы, хрусталика и стекловидного тела так, что фокус для разных участков спектра лежит на разном расстоянии от хрусталика, попадают на сетчатку, состоящую из одинаковых палочек и одинаковых колбочек. Чувствительные приёмники расположены так, что палочки воспринимают почти всю видимую область спектра (так как они длиннее колбочек), однако чувствительность их максимальна (в зависимости от интенсивности света и с учетом поглощения глазных сред) либо в сине-фиолетовой, либо в зелёной области (благодаря наличию в них светочувствительного пигмента родопсина). Колбочки воспринимают также почти всю область спектра, но преимущественно - жёлтую. Они имеют два участка, один обладает наибольшей чувствительностью в жёлто-зелёной области спектра (благодаря светочувствительному пигменту хлоролабу), другой — в оранжевой (благодаря светочувствительному пигменту эритролабу). Получается, что колбочка выдаёт несколько сигналов: один несет информацию об общей яркости, второй — о том, какой участок сильнее освещён и во сколько раз. Первый сигнал колбочки смешивается с сигналом палочки в нейроне, который формирует первый сигнал цветности Y и общий яркостный сигнал Z. Второй сигнал колбочки представляет собой второй сигнал цветности X. Все три сигнала (см. рис. 3) по нервным волокнам передаются в мозг, где сравниваются с памятью и вызывают ощущение цвета.
Эти процессы позволяют описать их математическими зависимостями, которые здесь не приводятся.
Так, как фотоответ рецептора имеет логарифмическую зависимость от освещённости, точка баланса сигналов отношения несколько смещается в зависимости от интегрального спектрального состава освещения, что обеспечивает постоянство цвета. Эта особенность известна под названием «цветовой адаптации».
Геометрическая интерпретация цвета[править | править код]
Цветокоординатная система

Если нанести в определённой последовательности все цвета воспринимаемые глазом на поверхность, они займут некую область, где каждому цвету будет соответствовать точка. Положение любой точки на плоскости можно определить всего двумя значениями координат, X и Y (в прямоугольной системе координат).
Несложно создать такую цветокоординатную систему. Известно, что белый цвет можно получить смешиванием всего двух цветов. Таких пар цветов может быть неограниченное количество. Причём, эти цвета могут быть спектральными, а могут и не быть таковыми. Логично принять точкой начала координат системы, точку соответствующую белому цвету. В этой точке цветовые сигналы равны нулю. Реально это будет точка серого цвета, зависящая от яркости, которая может изменятся от чёрного до белого (яркость это третья координата перпендикулярная нашей плоскости). Нанесём на нашу цветовую поверхность пары цветов взятых из непрерывного спектра, при смешивании дающие белый цвет. На нашей цветокоординатной системе они должны быть диаметрально противоположны относительно центра (белого цвета). Нанесём все пары цветов (образующих при смешении белый цвет) на нашу плоскость (при постоянной яркости). Получим некую область, описывающую положение спектральных цветов (непрерывного спектра полученного разложением призмой белого солнечного света) на координатной плоскости. Дополним нашу цветовую поверхность и неспектральными цветами например оттенками пурпурного являющимся противоположным оттенкам зелёного. Теперь проведём линии из точки начала координат (центр, белый цвет) в точки на обасти спектра с наиболее привычными нам цветами. Обнаружим закономерность: например линия жёлтый — синий проходит через центр нашей системы координат (белый). См. Рис. 4.
Кроме того заметим, что линия зелёный - белый, перпендикулярна линии жёлтый — синий. Если продлить линию зелёный — белый дальше, то она попадает на «пурпурный» цвет, который отлично воспринимает наш глаз, но которого нет в спектре. Примечательно, что пурпурный цвет на нашей системе расположен между красным и фиолетовым цветами, смешивая которые мы можем получить все оттенки пурпурного. Несложно заметить и проверить, что любой цвет на полученной плоскости можно получить смешиванием граничащих с ним цветов. Любой цвет воспринимаемый нашим глазом, можно легко выразить в этой системе вектором, идущим из точки начала координат (белого), в точку полученную геометрической суммой цветовых сигналов с координатой — Х (соотношение зелёного — пурпурного), координатой — У (соотношение синего — жёлтого) и сигнала яркости с координатой — Z.
Фактически мы получили цветовую координатную систему международной колориметрической системы МКО повторяющую всем известный цветовой круг. Кроме того мы нанесли координатную систему X, Y и Z.
Механизм перехода от дневного (цветного) зрения к сумеречному (монохромному)[править | править код]

На рис. 5 приведены кривые поглощения родопсина, пересчитанные с учетом поглощения его роговицей и хрусталиком. Восстановленный родопсин имеет только один максимум расположенный в области около 500 нм.
Разложенный родопсин также имеет один максимум, расположенный (с учетом поглощения роговицей и хрусталиком) в области около 465 нм. Это в случаях полной темноты и сильного освещения, во всех промежуточных условиях максимумов будет два [6]. Из этого следует, что могут быть такие условия, когда скорость разложения и скорость восстановления будут равны и оба максимума будут одинаковы по величине [7].
Согласно нелинейной модели дифференциально чувствительный элемент колбочка сравнивает спектральное распределение по двум длинам волн: соответствующим максимумам хлоролаба (540 нм.) и эритролаба (590 нм.). А дифференциальная система колбочка - палочка производит сравнение также по двум максимумам — суммарному максимуму колбочки, (570 нм.) и по «максимальному» максимуму родопсина, (или с синим — 465 нм., или с зелёным — 500 нм.).

Если освещённость высока, в качестве максимума сравнения используется «синий». Ширина воспринимаемой анализатором цвета области спектра велика, ощущаются все цвета. При уменьшении освещённости величина «синего» максимума уменьшается, появляется «зелёный» максимум, влияние которого состоит в увеличении чувствительности в зелёно-голубой области спектра. В силу уже рассмотренного нами свойства глаза корректировать изменение спектрального состава влияние «зелёного» максимума глазом замечено не будет. При снижении яркости влияние растущего «зелёного» максимума будет увеличиваться незаметно для глаза до того момента, когда оба максимума не станут близкими по величине. Эти условия, соответствующие наступлению сумерек, глаз уже не в состоянии скомпенсировать. Чувствительность голубой области спектра возрастает настолько, что белые объекты приобретают голубой оттенок. Это и есть известный издавна эффект Пуркинье, названный так именем чешского физиолога Яна Эвангелиста Пуркинье, впервые подробно описавшего его. Это единственное на сегодняшний день объективное объяснение этого эффекта.
При дальнейшем уменьшении освещённости скорость восстановления родопсина превысит скорость его разложения, «зелёный» максимум станет больше синего и система переключится на сравнение по «зелёному» максимуму. При этом, область спектра, воспринимаемая анализатором цвета (глазом), резко сузится. Спектральная характеристика глаза примет вид, изображенный на рис. 6. Цветовые сигналы станут ничтожно малыми и о распознавании цвета не может быть и речи. Глаз переключается на сумеречное (не цветовое) зрение. При этом кривая чувствительности глаза практически полностью совпадает с кривой ночного света.
Дефекты цветовосприятия.[править | править код]

Нелинейная модель цветовосприятия очень просто, наглядно и однозначно объясняет механизмы нарушения цветовосприятия глазом. Всего известны три частных случая цветоаномалии. Эти случаи наглядно показаны на рис. 7.
На рис. 7а, показана цветокоординатная система на которую нанесены длины волн разложенного солнечного спектра (кривая линия).
1.Отсутствует пигмент (сенсибилизатор), реагирующий на длинноволновую (жёлто-красную) область, — эритролаб. При этом модель описывает цвета воспринимаемые при поталогии дальтонизм 1-го рода — протанопия. Плоскость цвета (рис. 7а) вырождается из плоскости (сжимается) в прямую линию Yп, показанную на рис. 7б. На рисунке видно, что широкая часть спектрального состава вызывающего у нас ощущения от красного до зелёного цвета отображается на линии видения человека страдающего дальтонизмом первого рода (протанопа) на короткий отрезок линии Yп и поэтому воспринимается им как один цвет, в то время как широкая часть спектрального состава вызывающего у нас ощущения от зелёного до фиолетового цвета отображается на линии видения на большом протяжении и хорошо им различима.
2.Отсутствует пигмент реагирующий в основном на жёлто-зелёную область — хлоролаб. При этом плоскость цветовосприятия вырождается в линию Yд изображённую на рис. 7в. Такое цветовосприятие свойственно при дальтонизме 2-го рода — дейтеранопия. На рисунке видно, что широкая часть спектрального состава вызывающего у нас ощущения от красного до зелёного цвета отображается на линии видения человека страдающего дальтонизмом второго рода (дейтераноп) на длинный участок линии Yд и хорошо им различима, а широкая часть спектрального состава вызывающего у нас ощущения от зелёного до синего цвета отображается на линии видения на коротком отрезке и воспринимается им как один Цвет.
3.Отсутствует пигмент родопсин (в палочках) — так называемая «куриная слепота». При этом плоскость цветовосприятия вырождается в линию Xт (совпадающую с осью координат), нанесённую на рис. 7г. Этот случай и есть дальтонизм 3-го рода — тританопия. При этом видим, что участок спектра вызывающего у нас ощущения от красного до зелёного цвета отображается на тот-же отрезок линии видимости страдающего тританопией Yт, что участок спектра вызывающего у нас ощущения от зелёного до фиолетового цвета, т. е. он их не отличает.
Эти результаты полностью подтверждаются при исследовании цветоощущений людей которые подвержены этим типам дальтонизма. Других частных случаев при принятом принципе действия модели быть не может. В природе они тоже не наблюдаются. Если каких-либо пигментов меньше нормы, вырождение может быть не полным. Кроме аномалии цветовосприятия, модель может интерпретировать и три случая полной цветовой слепоты, но здесь мы на них останавливаться не будем, тем более, что и у людей они крайне редки.
Примечательно, что нелинейная теория зрения точно и чётко описывает как в случае тританопии человек воспринимает например радугу. В радуге цвета спектра расположены последовательно от фиолетового к красному. Но у тританопа плоскость воспринимаемого цвета вырождена в одну линию совпадающую с осью Х (см рис. 7г). Следовательно тританоп видит радугу (проекция кривой на ось Х) состоящую всего из двух цветов (назовём эти цвета, например "А" и "Б"). Но при этом края радуги (фиолетовый и красный цвет) он видит как цвет "А" (правый край проекции на ось Х), а к середине радуги цвет "А" плавно переходит в цвет "Б", через нейтральный серый (от правого края проекции на ось Х, к левому краю). В точке нейтрального цвета (пересечение с осью У) тританоп не отличает жёлтый и синий цвета от серого. Так как цветовые ощущения тританопа не совпадают с нормальным глазом, то мы не можем назвать цвета "А" и "Б" фиолетовым, красным, зелёным, жёлтым или ещё каким, они просто некие цвета, которые ощущает глаз при дальтонизме 3-го рода. Ни одна другая теория зрения не может дать однозначного объяснения специфики цветоощущения при дальтонизме.
Простейшая действующая модель глаза (колориметр)[править | править код]

На основе принципов сформулированных нелинейной теорией цветового зрения построена простая действующая фотоэлектрическая модель глаза, способная однозначно распознавать любые цвета и оттенки. Схема этой модели глаза показана на рис. 8. В качестве приёмников света используем три фоторезистора — два (R1, R2) в качестве сложного приёмника (колбочка) и один (R3) в качестве простого приёмника (палочка). Вместо пигментов воспользуемся светофильтрами (стеклянные фильтры не могут выцветать и восстанавливаться в зависимости от мощности излучения, так что переключаться на сумеречное зрение прибор не сможет, но для того, чтобы отличать цвета, это и не нужно). В качестве резисторов R1, R2, R3 — можно использовать фоторезисторы СФ2-1 или СФ2-2 из сульфида кадмия, в качестве резисторов R4, R5 — переменные резисторы СП-3 на 4,7 — 15кОм, V — диод (любой), включённый в прямом направлении (ограничивает ток через микроамперметр интенсиметра ИП3), Индикаторы ИП1 и ИП2 — микроамперметры с током полного отклонения 50 — 200 мкА с нулём посередине шкалы, Вык — выключатель, Е — источник питания напряжением 1 — 9 Вольт (в зависимости от чувствительности микроамперметров). R1 располагается за последовательно размещёнными фильтрами ОС-14 (оранжевым) и СЗС-21 (голубым), R2 размещён за фильтрами ОС-11 (оранжевым) и СЗС-8 (голубым), R3 размещаем за последовательно размещёнными фильтрами СС-5 (синим), СЗС-8 (голубым) и ЖС-3 (жёлтым). Фильтры СЗС-8 лучше использовать толщиной 4,5 мм, остальные 2 — 2,5 мм. При наладке схемы добьёмся одинакового ответного сигнала фоторезисторов R1 и R2 на дневной свет (нейтральный серый) параметры которого принимаем за начало координат. Устанавливаем переменный резистор R4 в среднее положение и прикрываем чёрной бумагой рабочую область более чувствительного фоторезистора ( R1 или R2) так, чтобы стрелка микроамперметра координаты Х была вблизи нуля. Аналогично поступаем с со вторым мостом R1 + R2 и R3 (резистор R3) при среднем положении R5 по прибору У. Перед началом работы переменными резисторами R4 и R5 приборы Х и У устанавливаются на нули при освещении фотоприёмников дневным светом.
Не смотря на то, что спектральные характеристики прибора несколько отличаются от характеристик глаза, он неплохо имитирует свойства нашего органа зрения. Так например, если из описанной действующей модели глаза исключить приёмник R1, моделирующий красночувствительную область колбочки с пигментом эритролаб, то он будет имитировать цветоаномалию первого рода — протанопию. При исключении приёмника R2, моделирующего зелёночувствительную область колбочки с пигментом хлоролаб, свойства колориметра будут описывать цветоаномалию второго рода — дейтеранопию. Исключение приёмника R3 моделирующего фоточувствительный пигмент родопсин в палочках, превратит прибор в цветоаномала третьего рода — тританопа.[8]. [9]
Этот прибор можно использовать во всех технологических процессах, где нужно быстро и точно определять и корректировать цвет.
Выводы[править | править код]
Нелинейная модель цветовосприятия созданная С. Ременко, просто и логично объясняет все свойства нашего зрения не ссылаясь на участие мозга, как это делают другие теории и гипотезы зрения. Модель глаза созданная на основе нелинейной теории рис. 8., очень проста и несмотря на это отлично работает в качестве колориметра. Любой воспринимаемый глазом цвет согласно этой модели элементарно просто записать в виде координат в декартовой системе.
Работы, проведённые в Институте прикладной физики и Институте химии АН МССР, совместно с кафедрой офтальмологии Кишинёвского Государственного медицинского Университета дали возможность предложить нелинейнyю модель анализа цвета, позволяющую представить цвет в виде физических параметров , являющихся результатом преобразования параметров излучения посредством известных физических законов и приборов.
Принцип модели основан на измерении отклонения точки цвета в цветовом теле от яркостной оси координат, вдоль которой лежат ахроматические тона.
Модель, представляющая аналог глаза, состоит из двух приемников, один из которых, дифференциальный — аналог колбочки — определяет отклонение точки цвета вдоль одной из осей координатной системы, а второй, с перестраиваемым максимумом спектральной чувствительности — аналог палочки — совместно с системой сравнения сигнала с суммарным сигналом первого — отклонение вдоль второй оси. Яркостный сигнал соответствует суммарному сигналу обоих приёмников. Такая модель представляет элементарную ячейку "одна колбочка — одна палочка" и позволяет описать и выразить физическими величинами все известные статические эффекты и аномалии цветового зрения. Макет колориметра, изготовленный на основе модели, позволяет с высоким быстродействием, определяемым быстродействием электронных элементов измерить физические параметры цвета в нелинейной координатной системе, свободной от многих недостатков системы МКО.
Физические законы, на которых основана модель , позволяют дать точные определения колориметрических терминов, отделить психологическую сторону, описать цветовые явления физическими параметрами и соотношениями[10].
Нелинейная модель позволяет дать четкие определения колориметрических характеристик излучения, основой которых являются физические закономерности, выражаемые математическим языком и величины которых можно измерить прибором. Предложены следующие определения:

Цвет — физический параметр, определяющий степень воздействия и спектральное распределение всех излучений, различимых фотоприемным устройством, характеристики которого соответствуют характеристикам глаза среднего наблюдателя.
Цветность — (Ц) векторная величина, определяющая спектральное распределение излучения.
Цветовой тон — (Ц тон) параметр, определяющий направление вектора цветности.
Яркость - (Я) координата Z.
Насыщенность — параметр излучения, определяемый отношением цветности (Ц) к интегральной яркости (Я) излучения.
Ахроматическое излучение — излучение, цветность которого равна нулю.
Противоположные цвета — цвета, векторная сумма цветностей которых равна нулю.
Метамерные цвета — цвета различного спектрального состава, векторы цвета которых равны.
Нелинейная система измерения цвета — система измерения цвета посредством трех взаимно независимых физических параметров: яркости излучения и двух функций отношения интенсивностей излучения в различных областях спектра.
Нелинейная цветокоординатная система — прямоугольная трехмерная координатная система бесконечной протяженности, описывающая параметры всех излучений, воспринимаемых фотоприёмным устройством, характеристики которого соответствуют характеристикам глаза среднего наблюдателя.
Координаты цвета — совокупность трех взаимно независимых параметров,определяющих положение вершины вектора цвета.
Отмеченные закономерности реализуются в устройстве, представляющем собой быстродействующий колориметр прямого отсчета, с помощью которого координаты цвета определяются по отклонению точки цвета от начала координат, лежащего на яркостной оси, вдоль двух выбранных разрезов. Характеристики колориметра определяются стандартизованными характеристиками "глаза среднего наблюдателя"[11].
Популярно о Нелинейной двухкомпонентной теории цветовосприятия[править | править код]
Совершенно не сложно популярно объяснить принцип цветовосприятия лежащий в основе работы нашего глаза.
Спектр оптического излучения — одномерный. Зачем использовать три датчика, для того, чтобы определить преобладающий цвет раздражителя? Ведь ни кому не придёт в голову измерять температуру тела тремя градусниками, а потом складывать, вычитать, делить и умножать, чтобы узнать истинную температуру! Кроме того, биологи сами себе противоречат когда утверждают, что клетка может складывать, вычитать, делить и умножать. Посудите сами: клетка питается, в ней накапливается определённый заряд. При внешнем раздражителе этот заряд выдаётся клеткой в виде электрического импульса фиксированной амплитуды. Если клетку чем либо быстро "возбудить" два раза подряд, не факт, что она обязательно выдаст два импульса. Клетке необходимо время для восстановления заряда. А как клетка может выполнить операцию умножения? В клетке нет кэш памяти и набора команд с инструкциями, что делать с тем или иным раздражителем. Зато любая клетка может выдать сигнал вызванный определённым соотношением нескольких внешних раздражителей.
Принцип работы глаза с позиции нелинейной теории цветовосприятия очень прост. Попроще объяснить его на примере работы обычных рычажных весов (например тех самых, что держит в одной руке Фемида — богиня правосудия: перекладина и две подвешенные к краям чашки). Пока, допустим, что эти весы подвешены не к руке статуи, а к ещё одним пружинным весам рис. 9а.

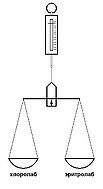
Рис. 9а. Модель поясняющая принцип работы колбочки глаза. Возбуждение хлоролаба уравновешено возбуждением эритролаба

Рис. 9б. Модель поясняющая принцип работы колбочки глаза. Возбуждение хлоролаба сильнее, чем возбуждение эритролаба

Рис. 9в. Модель поясняющая принцип работы колбочки глаза. Возбуждение хлоролаба слабее, чем возбуждение эритролаба



Давно открыто, известно и доказано, что в сетчатке глаза существуют два типа фоточувствительных рецепторов колбочки и палочки. В колбочке находятся одновременно два фоточувствительных пигмента эритролаб и хлоролаб[12]. Спектры их чувствительности перекрывают всю видимую область, перекрывают друг друга и отличаются только рядом расположенными максимумами чувствительности рис. 3а. Представим колбочку в виде весов на одной чашке которых хлоролаб, а на другой эритролаб рис. 9а. При воздействии на колбочку излучения длина волны которого находится как раз между максимумами чувствительности хлоролаба и эритролаба рис. 3а) (вызывает ощущение жёлтого цвета) раздражение хлоролаба и эритролаба будет одинаково, чаши весов уравновешены, но пружинные весы отмечают воздействие раздражителя — это и есть сигнал жёлтого цвета.
При воздействии на колбочку более коротковолнового излучения, относительно излучения вызывающего ощущение жёлтого цвета (в сторону синей области спектра), "чаша" хлоролаба будет тяжелее, чем "чаша" эритролаба рис. 9б. Рычажные весы покажут этот разбаланс, при этом и пружинные весы также покажут наличие сигнала раздражителя.
Если воздействовать на колбочку более длинноволновым излучением относительно излучения вызывающего ощущение жёлтого цвета (в сторону красной области спектра), "чаша" хлоролаба будет легче, чем "чаша" эритролаба рис. 9в. Рычажные весы так-же покажут этот разбаланс (уже в другую сторону), а пружинные весы также покажут наличие сигнала раздражителя. По уровню этого разбаланса (отношению реакции эритролаба к хлоролабу) "рычажных" весов всегда можно точно определить длину волны раздражителя или преобладающую область спектра раздражителя. Что уникально, так это то, что результирующие данные по цвету совершенно не зависят от уровня освещения!, чем и достигается такой широчайший рабочий диапазон работы глаза. Эту информацию и выдаёт клетка в виде импульсов.

Выше было показано, что весы прекрасно иллюстрируют принцип работы колбочки. Равновесие чашек весов, при воздействии света, показывает на преобладание длины волны вызывающей ощущение жёлтого цвета. По направлению и углу отклонения чашек весов мы можем точно определить любую длину волны (или преобладающие длины волн) воздействующие на колбочку от крайнего фиолетового, до другого края — красного. Однако в палитру цветов воспринимаемых нашим глазом входят также цвета не существующие в спектре. Это не удивительно, ведь в нашем глазу помимо колбочки есть ещё один тип рецепторов палочка. В палочке содержится фоточувствительный пигмент родопсин. При достаточном освещении (дневном зрении) родопсин разлагается и при этом максимум его спектра поглощения приходится на синюю область. Достроим нашу модель глаза.
Рычажные весы имитирующие работу колбочки, подвесим к одной из чашек других рычажных весов. Вторую же чашку, вторых весов, будет играть палочка, реагирующая на коротковолновую (синюю) область спектра (колбочки слабо чувствительны в синей области спектра). А уже эти "вторые весы" подвешены к пружинным весам, которые "держит богиня правосудия Фемида" рис. 10. Что мы поучили? Мы получили действующую модель глаза! Первые рычажные весы (хлоролаб — эритролаб) имитируют работу колбочки, показывая координату цвета на линии зелёный — пурпурный. При этом если колбочку осветить излучением вызывающим ощущение жёлтого цвета, то сигнал (вес) будет, но первые весы будут в равновесии. Зато левая чашка вторых весов станет тяжелее и перевесит правую чашку — палочку (датчик синего цвета). Получается, что вторые весы показывают положение координаты цвета на линии жёлтый — синий.
Теперь посмотрим на всем известный цветовой круг рис 11. На нём два противоположных цвета (относительно центра круга) в смеси всегда дают серый (от белого до чёрного в зависимости от интенсивности).

Нанесём на наш цветовой круг две прямые линии, первую линию (первые весы — колбочка) через точки: зелёный - центр круга — пурпурный (колбочки, как и любые клетки живого организма чувствительны к ультрафиолетовой и фиолетовой областям спектра, поэтому при широкополосном спектре освещения мы видим не красный цвет, а пурпурный) и вторую линию (вторые весы — "взвешивающие" отношение сигнала колбочки к палочке) через точки: жёлтый — центр круга — синий. И что мы получили? Мы получили, что эти две линии пересекаются в центре круга и они ПЕРПЕНДИКУЛЯРНЫ! Вот вам готовая, удобная, понятная Декартова система координат цветового пространства. Любой цвет на этом пространстве выражен однозначным соотношением сигнала от эритролаба — хлоролаба в колбочке (линия зелёный — пурпурный (координата "Х")) и взвешенного сигнала колбочки к палочке (линия жёлтый — синий (координата "У"). Пружинные весы к которым подвешена эта система показывают суммарную "тяжесть" полного сигнала, т. е. его ЯРКОСТЬ (координата "Z") рис 12.
Вот и всё! Для полноценного восприятия всех возможных цветов и оттенков глазу не нужны три различных типа колбочек, мнимый пигмент цианолаб и "высшая нервная деятельность мозга". Результат оценки цвета ОДНОЗНАЧНО выдаёт пара рецепторов колбочка-палочка. Это очень просто, надёжно и очень рационально. Любой цвет можно выразить тремя координатами в декартовой системе координат цветового пространства. Для нормальной работы глаза не нужны три типа колбочек и на лицо принцип оппонентности, дающий объяснение многим свойствам зрения. Фактически сама система колбочка — палочка производит полный анализ спектра раздражителя и выдаёт уже готовый цветовой сигнал без какого либо участия мозга.
Именно поэтому сторонники трёхкомпонентных теорий, так до сих пор и не могут найти: ни мнимого фотопигмента "цианолаба", ни трёх типов колбочек, ни трёх типов нервных окончаний по которым идут сигналы от "красной", "синей" и "зелёной" колбочек, ни других атрибутов, необходимых для трёхкомпонентных теорий. Их просто не существует. Зато того, что известно и открыто в строении глаза на сегодняшний день, более чем достаточно для полного описания функционирования глаза согласно Нелинейной Двухкомпонентной Теории Цветовосприятия.
Первый в мире действующий макет глаза (создан в 1975 году)[править | править код]

Фото 1. Внешний вид прибора

Фото 2. Внешний вид сенсора

Фото 3. Внутреннее устройство прибора



Данный действующий макет глаза был создан автором нелинейной двухкомпонентной теории цветового зрения С. Ременко в 1975 году. Этот макет работает на принципах заложенных в основу нелинейной теории цветовосприятия. Схема макета показана на Рис. 8. Уникальность прибора состоит в том, что в нем отсутствуют какие либо активные электронные компоненты (транзисторы, усилители, компараторы, процессоры и пр.), при этом макет полностью моделирует работу глаза, однозначно определяя физическую координату любого существующего цвета в трёхмерной декартовой системе координат Рис. 12.
Внешний вид прибора показан на фото 1.
1. Левый индикатор показывает координату цвета на оси Х (зелёный — пурпурный).
2. Правый индикатор показывает координату цвета на оси У (жёлтый — синий).
3. Средний индикатор показывает уровень яркости — координата Z.
Любой цвет однозначно интерпретируется этими тремя координатами.
Внизу слева расположен разъём для подключения сенсора моделирующего работу колбочки и палочки глаза. Далее расположен 5-ти позиционный переключатель имеющий следующие режимы:
1. О — отключен.
2. Н — нормальное зрение.
3. I — режим моделирующий цветовосприятие при дальтонизме 1-го рода — протанопия (Отсутствует пигмент эритролаб в колбочке).
4. II — режим моделирующий цветовосприятие при дальтонизме 2-го рода — дейтеранопия (Отсутствует пигмент хлоролаб в колбочке).
5. III — режим моделирующий цветовосприятие при дальтонизме 3-го рода — тританопия (Отсутствует пигмент родопсин в палочке).
Далее расположены три потенциометра которыми выставляются "нулевые параметры" на индикаторах при направлении сенсора на объект (источник) или освещении сенсора источником широкополосного "белого" цвета соответственно по осям: Х (зелёный — пурпурный), У (жёлтый — синий) и Z (яркость).
На фото 2 показан внешний вид сенсора содержащий аналог колбочки и палочки (три фоторезистора размещённые под светофильтрами соответствующими спектрам чувствительности фотопигментов родопсина (содержащегося в палочке), хлоролаба и эритролаба (содержащихся в колбочке).
На фото 3 показано внутреннее строение макета. Слева внизу видны два элемента АА питающие макет.
p. s. всё гениальное — просто.
Ссылки[править | править код]
- ↑ Дж. Уолд. Scientific American, Восприятие, механизмы и модели, Москва, издательство «Мир», 1974 год, 138.
- ↑ Ричард Филлипс Фейнман. Фейнмановские Лекции по Физике, том № 3, гл. 35 (Цветовое зрение), стр. 175.
- ↑ Хьюбел Дэвид. Глаз, мозг, зрение, Биология, Москва, издательство «Мир», 1990 год.
- ↑ С. Ременко, «Цвет и зрение», «Картеа Молдовеняскэ», Кишинёв, 1982 г.
- ↑ «Visual Pigments of Single Primate Cones» W. B. Marks, W. H. Dobelle, E. F. Mak Nichol Department of Biophysics, Johns Hopkins University, Baltimore, Maryland 21218. Science, Vol. 143, p. 1182, 1964.
- ↑ Островский М. А., Федорович С. Е., Голубев И. Н., 1967, Биофизика, 12 : 877.
- ↑ Hubbard R., Bownds D., Yoshizawa T., 1965. Cold Spring Harbor Symp. Biol., 30 : 301.
- ↑ С. Ременко, «Цвет и зрение», «Картеа Молдовеняскэ», Кишинёв, 1982 г.
- ↑ С. Ременко, «Как построить глаз», журнал «Техника Молодёжи», 1981 год,№ 7, стр. 21 — 25.
- ↑ С. Д. Ременко, «Нелинейная модель измерения цвета и уточнение терминов колориметрии», Всеакадемический семинар по проблемам стандартизации и метрологии, Ташкент, 20 — 25 ноября 1986 год, стр 41 — 42.
- ↑ С. Д. Ременко, «Определение основных понятий в области колориметрии и измерения цветовых параметров излучения», V Всеакадемический семинар по проблемам стандартизации и метрологии Ереван, 16 — 20 ноября 1987 год, стр 58 — 59.
- ↑ «Visual Pigments of Single Primate Cones» W. B. Marks, W. H. Dobelle, E. F. Mak Nichol Department of Biophysics, Johns Hopkins University, Baltimore, Maryland 21218. Science, Vol. 143, p. 1182, 1964.
См. также[править | править код]
- Теории цветового зрения
- Зрительная система
- Зрение человека
- Зрение
- Метамерия (цвет)
- Опровержение трёхкомпонентной гипотезы цветного зрения
- Иллюзии и парадоксы цветового зрения
- родопсин
- Сетчатка
- Палочки (сетчатка)
- Колбочки (сетчатка)
- эффект Пуркинье
- Дальтонизм