Сравнительная таблица трёхкомпонентной и нелинейной теорий цветного зрения
Сравнение теорий цветного зрения трёхкомпонентной и нелинейной обсуждается с точек зрения трихроматизма и нелинейеой теории зрения. С точки зрения трёхкомпонентного принципа цветеого зрения материал рассматривается в основном на базе исследований на живых клетках, исследований человека, опыты на животных также служат материалом для сравнений. Все данные материала представлены в виде таблиц сравнений.
| 1 Явление или факт | 2А Пояснение, с позиции 3-х комп. модели и следствия | 2Б Критика этого пояснения: с т.з. нелинейной модели | 3А Пояснение с позиции нелинейной модели, её предсказания и следствия | 3Б Критика этого пояснения: с т.з. 3-х компонентной модели | 4 Обсуждение и сравнение |
|---|---|---|---|---|---|
| 1. В глазу обнаружено только 3 типа фоторецепторов палочки и колбочки и ганглиозных фоторецепторов ipRGC. | 1.2А Доказано, что в глазу в фокальной поверхности сетчатки существуют палочки и 3-и (или даже 4-ре) типа колбочек (См. Цветное зрение у птиц (версия Миг), Труды доктора Р.Е.Марка и его лаборатории, Ретиномоторная реакция фоторецепторов (версия Миг) и др.), а также недавно открытые фоторецепторы ганглиозного слоя сетчатки ipRGC, которые синапстически связаны с колбочками и палочками и головным мозгом. Фоторецепторы ipRGC чувствительны к синим лучам света, содержа фотопигмент меланопсин (версия Миг). Меланопсин реагирует на ультрафиолетовые и синие лучи света. | 1.2Б Ни каких различий между существующими колбочками сетчатки не существует. | 1.3А Для цветовосприятия достаточно только одного типа палочек и одного типа колбочек | 1.3Б С точки зрения ретиномоторной реакции фоторецепторов колбочки работают в условиях дневного освещения выше 498нм (цветное зрение) и палочки — в условиях сумеречного освещения до 498нм (не цветное зрение), при этом днём только колбочки, вечером — только палочки (граница 498нм). Снимки срезов сетчатки при флюоресцентной микроскопии фиксируют колбочки S,M,L (синие, зелёные, красные). Колбочки-S, работающие в синей части спектра света с длиной волны свыше 498нм, аналогично
— палочкам, но в зоне — меньше, чем 498нм. Откуда при дневном зрении участвуют только три одинаковые колбочки, которые оппонентно формируют выходные нормализованные сигналы в мозг S,L,M, то есть синего, зелёного и красного цветов, при восприятии лучей дневного света как отражённых так и прямых в составе пучка основных лучей КЗС — в условиях трёхкомпонентного цветного зрения. (См. Труды доктора Р.Е.Марка и его лаборатории, Теория трёхкомпонентного цветного зрения (версия Миг). |
(Обс.1.4) В глазу обнаружено 3 типа фоторецепторов палочки, колбочки и фоторецепторов ipRGC ганглиозного слоя сетчатки (Фоторецепторы ipRGC обнаружены недавно).
В то время, как с точки зрения нелинейной теории зрения в глазу при цветном зрении работают блоки колбочка+палочка, но доказано, что при цветном зрении (дневное осаещение) работают только колбочки. (См. Ретиномоторная реакция фоторецепторов (версия Миг)). |
| 2. Объяснение механизма различных типов дальтонизма | 2.2А Согласно последним данным действующей и расширяющейся трёхкомпонентной теории цветного зрения объясняются дефекты цветовосприятия глаза. На сегодняшний день тщательно описаны три типа цветоаномалии:
|
2.2Б Трёхкомпонентные теории в принципе не могут точно описать механизм цветоаномалий глаза | 2.3А Отлично описывает механизм всех известных цветоаномалий. Действующая модель глаза, построенная на основе теории, моделирует все известные виды цветоаномалий глаза. | 2.3Б Модель глаза, построенная на основе колориметра на основе фотодиодов-фототранзисторов вообще не способна фототрансдуцировать сигнал предметной точки, т.к нет принципа оппонентного отбора цвета с участием фоторецептора ipRGC и мозга на рецепторном уровне (не цветового) с дальнейшим его прохождением в мозг, где формируется оптическое изображение в цвете, где в процесс на основе трёх отобранных сигналов S,M,L предметной точки идёт формирование одного общего сигнала предметной точки оптического изображения в мозгу. При это мозг работает по типу АЦП как в цифровой фотографии. То есть мы у себя создаём аналоговое оптическое изображение в сознании. Данный процесс пока непостижим. Все цвета у нас индивидуальные, которые можно только сравнить при опросе. С точки зрения нелинейной теории зрения все получаемые цвета никакого отношения к цветам человека не имеют. Их можно только подогнать и сравнить с эталонным среднестастическим цветом при опросе большой группы участвующих людей. Откуда, как можно говорить об аномалиях и других заболеваниях глаза, не видя сами живые клетки, сам глаз. | (Обс.2.4) См.2.2А |
| 3.Фоточувствительные пигменты сетчатки.
Описаны три типа разновидностей фотопигмента колбочек при цветном зрении Йодопсина: Эритролаб, Хлоролаб и ещё не выделенный фотопигмент Цианолаб, работающих только в условиях дневного освещения. При сумеречном, не цветном зрении работают только палочки, содержащие фотопигмент Родопсин (версия Миг). В ганглиозных клетках сетчатки фоторецепторах ipRGC, участвующих в зрительном процессе, также содержится фотопигмент меланопсин (версия Миг), чувствительный к части спектра фиолетово-синих лучей. |
3.2А С точки зрения принципа трихроматизма при цветном зрении работают одинаковые колбочки, выделяющие фоточувствительные фотопигменты Йодопсин (версия Миг)а во внешнюю мембрану в зависимост от сфокусированного на неё луча части спектра: (красного)Эритролаб , (зелёного) Хлоролаб и ещё не выделенный, но индентифицированный фотопигмент синего цвета цианолаб. При сумеречном, не цветном зрении, работают палочки, содержащие фотопигмент Родопсин (версия Миг). При этом любая колбочка блока из трёх или четырех колбочек реагирует на спроектированный на неё луч предметной точки и в зависимости от отобранного оппонентно более яркого сигнала внешняя мембрана колбочки трансдуцирует этот сигнал в мозг. Например, выделенный оппонентно красный луч предметной точки (из кружка нерезкости) распознаётся выделяющимся фотопигментом эритролабом и далее направляется по своим каналам в мозг. Выделенный сигнал на данном рецепторном уровне не цветной. Трансформированный, а может и усиленный, он попадает в мозг. Аналогично остальные колбочки блока трансформирующие зелёный луч, который распознаётся фотопигментом хлоролабом и синие сигналы этой же точки распознаются пока не открытым, но индетифицированным фотопигментом синего цвета цианолабом, которые также отправляются в мозг (т.е. эквивалентные биосигналы КЗС).
(Примерно так заимствована у глаза схема работы цветной фотографии). На этапе оппонентного отбора цвета в этом процессе участвуют все возможные сочетания цветов спроектированной конкретной предметной точки на внешнюю мембрану колбочки. Отсюда не может и быть речи о получении конечного сигнала по типу биологической системы получения оптического изображения в мозгу при помощи фотоаппарата или модели глаза (колориметра). Наше сознание, головной мозг, глаз с сетчаткой — это биологическая оптическая система, которую создала природа, она еще не постижима. Человек многое познал, перенял, чтобы помочь при недугах. Например, бионический глаз. И то, что ещё не открыт фотопигмент цианолаб, то это дело времени. А выделенная колбочка-S (синяя), вообще подтверждает трёхкомпонентную теорию цветного зрения, где работают три вида колбочек S,M,L. (См. Лаборатория Р.Е.Марка (версия Миг), Денситометрия и фотопигменты колбочек, Индентификация разновидностей йодопсина — цианолаба). |
3.2Б требуется наличие так и не найденного до сих пор пигмента цианолаба. Кроме того спектры поглощения эритролаба и хлоролаба (см.) совершенно не соответствуют основным цветам Поэтому постулирование трёхкомпонентной теории о «смешивании» принципиально не позволяет получить всю гамму видимых глазом оттенков. | 3.3А Колбочка со смесью фотопигментов: эритролаб и цианолаб (входят в состав колбочкового пигмента Йодопсин), является нелинейным датчиком однозначно показывающим преобладающее воздействие смеси любых спектров видимого света в форме определённого цвета. Определение цвета происходит путём сравнения (взвешивания) фотоответов эритролаба и цианолаба. | 3.3Б Согласно ретиномоторной реакции фоторецепторов только одни колбочки работают при дневном освещении, обеспечивая цветное зрение в условиях освещения лучами части спектра длиной волны более 498 нм, палочки работают отдельно в условиях сумеречного освещения (не цветное зрение) при освщении лучами части спектра длиной волны менее 498 нм. Колбочки работают с выделением разновидностей фотопигмепа йодопсина во внешней мембране колбочек, палочки работают с содержанием единственного фотопигмента в мембране родопсина. При этом при оппонентном отборе (анализе) наиболее ярких сигналов предметной точки основных лучей (адэкватных электромагнитных волн) КЗС на рецепторном уровне они посылаются в мозг. В мозгу на нейронном уровне (синтез) эти КЗС на базе аддитивного синтеза ("смешения" адэкватных электромагнитных волн) оппонентно-отобранных лучей КЗС формируются аналоговые цветовые точки цветного оптического изображения. Таким образом, в зрительной коре и происходит процесс аддитивного смешивания КЗС. Этот биохимический процесс цветного зрения отличается от колориметрии при составлении атласов цветов, где все видимые световые лучи создаются искусственно, без оппонентного отбора с применением специальных излучателей видимых световых лучей и смешивание этих монолучей производят также при помощи спецприборов. Получаемые оттенки оцениваются среднестатистическим наблюдателем. Смотри Ретиномоторная реакция фоторецепторов (версия Миг), Цветное зрение у человека (версия Миг), Труды доктора Р.Е.Марка и его лаборатории, Денситометрия и фотопигменты колбочек, Индентификация разновидностей йодопсина — цианолаба). | (Обс.3.4)
|
| 4.Опсины (версия Миг) (Фотопигменты сечаткти газа) | Обс.4.2А Доказано на исследованиях живой клетки (но не на фототранзисторах!)[1], что опсин как фотопигмент имеет пять групп белков-опсинов, которые принимают участие в процессе зрительного восприятия на рецепторном уровне с превращением света в выделяемый электрохимический сигнал, равный значениям основных цветов КЗС (не цветной), и являются первым этапом (рецепторным) в каскаде зрительной фототрансдукции. В целом эти опсины можно разделить на две группы:
Разновидности опсина йодопсин (версия Миг) содержится в колбочках, которые в зависимости от падающего на их внешнюю мембрану луча предметной точки, выделяют фотопигменты: Эритролаб(красный цвет),Хлоролаб (зелёный цвет), еще не выделенный цианолаб (синий цвет). Однако, выделение синего пигмента типа цианолаба в колбочках доказано при микроскопии сетчатки c получением изображения в плане колбочки-S синего цвета (его название при выделении может быть разным). Фотопигмент синего цвета есть у колбочек, как и у палочек родопсин (Менее 498нм). Отличие в том, что полочки работают в зоне спектра до 498нм, колбочки — более 498нм; следует полагать, что фотопигмент цианолаб более чувствителен, молоинерционный и обеспечивает колбочкам восприятие цветовых лучей более длинноволновых. (Более 498нм) |
4.2Б Нет ни одного доказательства связи, что один опсин всегда соответствует только одному фотопигменту, и в свою очередь тому, что в каждой колбочке содержится только один (свой) пигмент. | 4.3А Опсины (версия Миг) входят в состав фоточувствительных пигментов (эритролаб, хлоролаб, родопсин (версия Миг)). В состав некоторых из существующих фоточувствительных пигментов входит сразу несколько типов опсинов. | 4.3Б Опсины (версия Миг) входят в состав видоизменяющихся фоточувствительных пигментов колбочек, которые находятся у каждой колбочки. В зависимости от поглощённого луча, выделяется нужный вид опсина, который этот луч обрабатывает и выделяет в сигнал для передачи в мозг. Например, синий луч обрабатывается выделяющимся видом опсина, чувствительного к сине-фиолетовому спектру лучей с длиной волны более 498нм цианолабом. (См. Йодопсин (версия Миг),Индентификация разновидностей йодопсина — цианолаба, Денситометрия и фотопигменты колбочек). | Обс.4.4 См. Опсины (версия Миг) |
| 5. Эффекты Бетцольда-Эбнея и Бетцольда-Брюкке, Эффект Пуркинье, сумеречное зрение и пр. | 5.2А Чтобы разобраться в парадоксах, иллюзиях прежде всего нужен определённый уровень знаний с постановкой задачи и проведением эксперимента замеченного парадоксального события на приборах, позволяющих рассмотреть все участвующие факторы в их динамике. Например, чтобы оценить все парадоксы зритедьной системы нужно исследовать живой глаз и всё, что с ним связано. Что является основой трихроматизма. Вся сложность и состоит в том, что мы не можем пока увидеть, или изучить сознание, которое обобщает созданные двойные оптические изображения в мозгу на рецепторном и нейронном уровнях одновременно или в динамике, каскадом, в виде видео. Именно здесь вступает в силу работа сознания и мозга, которое выносит окончательный иллюзорный вердикт.
Поэтому, чтобы помочь работе мозга, для устранения иллюзий, воздействующую информацию человек разделяет на части, которые он предварительно анализирует с использованием имеющихся методик, средств измерения, например, в метрологии — метра, при помощи кино — видео. Метром можно замерить длину каждого из двух одновременно полученных изображений в мозгу, выданных для сравнения. (Как в цирке). Это можно оценить, когда мы измерим длину каждого предмета. При видео можно в замедленной форме показать работу каскада событий в динамике, в расчленённой замедленной форме. (Например в спорте, при тренировочном процессе). Здесь например, можно это проанализировать с точки зрения возможностей зрительного процесса трихроматизма и нелинейной теории цветного зрения. Во-первых, полноценную инфрмацию мы можем получить при дневном освещении, когда работают только колбочи (палочкив в это время спрятаны и находятся вне зоны попадония лучей предметных точек). Днём проходит вся наша жизнь в цвете, именно когда выдвинуты как грибы колбочки-S,M,L, которые являются основой выдачи любой полноценной инфомации в мозг. Откуда, информация любая, в том числе и исследовательская не должна создавать иллюзии. Например, при исследовании зрительного процесса с получением и выдачей не проверенной информации в мозг (например, в современных условиях без применения флюоресцентной микроскопии живой ткани), или с другой стороны — выдачей в мозг многочисленных графиков, кривых, полученных при исследовании работы роботов, вместо живого глаза и мозга, при которых не учитывается работа клеток мозга и самое главное, работа с получением информации не совпадающей с имеющейся или отсутствующей для сравнения с объективно имеющейся. Например, получаем парадокс, который мы наблюдаем при создании и попытке внедрения в наше сознание нелинейной теории цветного зрения, где выводится, что в основе цветного зрения работает палочка, вместо колбочки-S. (Хотя доказано, что палочка не участвует в работе зрительной системы при дневном освещении, что может служить примером создания иллюзий в нашем сознании). (См. Ретиномоторная реакция фоторецепторов (версия Миг)). Возьмите нанотехнологию. Здесь иллюзии вообще исключаются. Только на базе современных микроскопов с огромным разрешением научились рассматривать, захватывать и перемещать атомы и молекулы вещества, создавая предсказуемые и непредсказуемые совершенно ранее не типичные физические свойства одного и того же материала. Применение флюоресцентных микроскопов в исследованиях живой сетчатки, позволяют нам рассмотреть сетчатку со всеми клетками на атомно-молекулярном уровне в режиме стерео. Откуда применяемые методики исследований в трёхкомпоннтной теории цветного зрения исключают выдачу иллюзорной информации в мозг, где для анализа и сравнения всех событий в области цветного и вообще зрения мы получаем объективную информацию, которая не порождает парадоксы и иллюзии в нашем сознании, дает возможность продвигаться вперёд в изучении существующих не познанных ялений, парадоксов и иллюзий зрительного процесса. |
5.2Б Трёхкомпонентная теория даже приблизительно не может дать однозначного объяснения этих эффектов | 5.3А Отлично описывает механизм эффектов. Действующая модель глаза, построенная на основе Н.теории, моделирует перечисленные эффекты и вообще все известные эффекты работы глаза. | 5.3Б См.5.2А, Проблемы современных теорий цветового зрения (версия Миг) | 5.4 См.5.2А |
Выводы[править | править код]
Анализ данных цветовосприятия в последние 35‒40 лет показывает, что c появлением новой техники, технологии исследований, учёным удалось доказать, что принцип трихроматизма (принцип трёхкомпонентного цветного зрения) является основой цветного зрения. На базе исследований на живых клетках при воздействии на них видимых лучей света (цвета) в цвете удалось рассмотреть колбочки S,M,L (RGB), которые участвуют в процессе дневного зрения, в процессе формирования цветного изображения в зрительных отделах головного мозга в трёхмерном пространстве в цвете, в стерео. C открытием третьего фоторецептора ганглиозного слоя сетчатки (см. ipRGC (~2007 год)), который расположен не в фокальной поверхности сетчатки, синаптически связан с колбочками, палочками, мозгом. Он выполняет важные функции при цветовосприятии в регулировании работы колбочек при восприятии спектра коротковолновых лучей (синих-фиолетовых), пропуская нормальные лучи спектра с длиной волны более 498нм, перенаправляя их в зону восприятия синих лучей колбочек-S, блокируя сильнодействующие синие лучи с высокой частой колебания — длинами волн менее 498нм. Он содержит оптический пигмент меланопсин (версия Миг), чувствительный к синим лучам (S) света.
Проводились исследования учёными Е. О. Загальской и В. П. Гнюбкиной из Института Биологии Моря ДВО РАН (2006 год), Владивосток. Цель этого исследования состояла в том, чтобы проанализировать ретиномоторные движения у взрослых рыб (zebrafish) и процесс созревания ретиномоторной реакции в его развитии у молодняка рыб. Показано, что ретиномоторные движения используются как адаптация зрения в условиях тёмно/светового освещения, показан механизм адаптации у рыбы-зебры. У взрослых особей гранулы меланоцитов.

В 2006 [3] году при помощи трансмиссионной электронной микроскопии пигментного слоя сетчатки исследован эпителий клеток сетчатки глаза, где вырабатывается пигмент меланин (коричневого и чёрного цвета), который обеспечивает открытие и закрытие зон освещения колбочек и палочек в зависимости от вида освещения. При дневном освещении данный пигмент блокирует работу палочек, которые ко всему ещё уходят в зону под колбочки. И наоборот, при сумеречном освещении палочки выходят из под прикрытия зоны прохождения синих и ультрафиолетовых лучей меланином, а колбочки опускаются в зону ниже палочек и находятся под прикрытием меланина. (См. рис. C). В конечном итоге подтверждено открытие явления ретиномоторной реакции фоторецепторов , работы колбочек и палочек в условиях дневного и ночного освещения учёными Е. О. Загальской и В. П. Гнюбкиной из Института Биологии Моря ДВО РАН, Владивосток.
- Рентгеноскопия сетчатки и отделов мозга, проведенная ещё в 1966, повторно в 2009 годах Р. Е. Марком, на атомно-молекулярном уровне подтверждает ранее сформулированые гипотезы о работе сетчатки, где заложены фоторецепторы колбочки, выделющие основные лучи спектра S,M,L («синие», «зелёные», «красные») (см. Фиг. R,рис. S), которые на рецепторном уровне воспринимают, трансдукцируют основные сигналы RGB оптического изображения (контурного, рецепторного) перед передачей их в высшие зрительные отделы (нейронные) головного мозга (см.Лаборатория Р.Е.Марка (версия Миг)). При этом следует подчеркнуть, что структукрно колбочки устроены одинаково, но в зависимости от места расположения они имеют разные морфологические размеры, например, в центральной ямке сетчатки они самые маленькие, так как плотно упакованы и при этом, в зоне ямочки диаметром до 1 мм расположены только колбочки красные и зелёные, где нет палочек вообще. Каждая колбочка способна оппонентно выделить один из основных цветовых сигналов S,M,L, КЗС, так как фотопигмент опсин в зависимости от сфокусированного и выделенного основного луча предметной точи способен в результате мутации белка пигмента принимать красную, зелёную и синюю окраску!
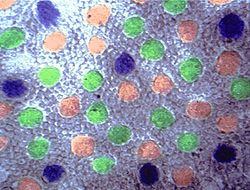 Рис. S. Распределение рецепторов в сетчатке бабуина . Синие колбочки в периферийной зоне ямки фовея с углом охвата 7-8 градусов были распределены регулярно, красные и зеленые колбочки были распределены беспорядочно всюду. Плотность распределения зелёных колбочек больше, чем красных, больше чем синих.[4]
Рис. S. Распределение рецепторов в сетчатке бабуина . Синие колбочки в периферийной зоне ямки фовея с углом охвата 7-8 градусов были распределены регулярно, красные и зеленые колбочки были распределены беспорядочно всюду. Плотность распределения зелёных колбочек больше, чем красных, больше чем синих.[4]

В настоящее время уже вообще не стоит вопрос о восприятии цвета в сетчатке блоками из трёх колбочек S,M,L (R.Marc, 1977).[4][5] Если говорить о восприятии синих монолучей синими колбочками, то уже в 1977 году получены легко рентгеновские снимки в плане фокальной поверхности сетчатки синей колбочки в ряду основных колбочек зелёных и красных (см. рис. S).
Группа учёных Лаборатории R.Marcа (Труды доктора Р.Е.Марка и его лаборатории) уже работают над созданием атласа основных нервных клеток сетчатки и отделов головного мозга, на основании электронной микроскопии их у кроликов и крыс. При этом используется разработаная мощная программа обработки данных современных микроскопов и компьютерных систем. При этом открывается возможность визуально анализировать работу здоровых и больных клеток с целью лечения целого ряда болезней (слепота, дальтонизм и др.[4]
Что важно, на рентгеноскопии среза сетчатки (cм. Фиг. R) видно, что основные фоторецепторы колбочки и палочки в процессе формирования оптического изображения работают независимо. Так синяя колбочка при дневном зрении воспринимает диапазон синих лучей под присмотром фоторецепторов ipRGC. Например, сильные сигналы ультрафиолетовых, синих лучей под действием ipRGC и мозга закрывают вообще ресницами глаза глаз от их попадания во внутрь (скорость реакции 1/2000сек) и имеются другие функции у них, связанные с трансдукцией синего сигнала.
- Доводы о том, что палочки и колбочки одинаковы и не имеют различий голословны и оторваны от жизни. В настоящее время получены изображения разных типов колбочек и палочек, которые отличаются мрфологически и функционально. (См. рис. 11). На приведенных изображениях колбочек птиц и черепах ещё раз показан вид колбочек, которые морфологически одинаковы, но различаются видом, в зависимости от места расположения и меняющемуся виду при работе. В глазу птиц даже есть двуглавые колбочки, расположенные в периферийной части и управляющие движением птиц.
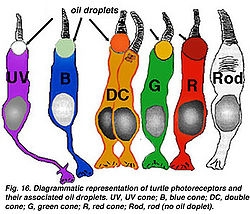 Рис.11;Блок колбочек черепахи с цветными жировыми капельками[6]
Рис.11;Блок колбочек черепахи с цветными жировыми капельками[6]

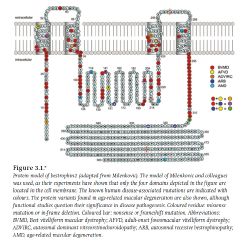 Рис. П; Мутация белка протеина (содержащего разновидность пигмента родопсин) в мембране колбочки человека в четырёх областяхЧто касается фотопигментов, то следует знать, что пигмент, например, фоторецептора или изменяют своё устройство или претерпевают уменьшение своего изображения (вида), когда они поглощают фотон. Это изменение происходит в устройстве или в окислительно-восстановительном процессе хромофора и выражается в изменении устройства белка или его деятельности, и вызывает каскад трансдукции сигнала. Примеры для пигментов фоторецепторов, находящиеся в сетчатке глаза: например, родопсин (версия Миг) (относящийся к сложным белкам хромопротеинам) вызывает жёлтый цвет; например, криптохром (cryptochrome), и билин (bilin) вызывает голубой цвет растений. Относительно фотопигмента цианолаба, который ещё не выделен учёными, то в результате фототрансдукции сигнала, пигмент, который светится в жировых капельках синим цветом, увиден и его можно назвать любым именем, важен цвет, который виден при исследованиях на снимках, и который имеется на синих колбочках-S. Не исключено, что он близок к меланопсину, билину. В любом случае, он является разновидностью фотопигмента опсина (См. Белок и передача зрительных биосигналов (версия Миг)) [цитата, необходимая]
Рис. П; Мутация белка протеина (содержащего разновидность пигмента родопсин) в мембране колбочки человека в четырёх областяхЧто касается фотопигментов, то следует знать, что пигмент, например, фоторецептора или изменяют своё устройство или претерпевают уменьшение своего изображения (вида), когда они поглощают фотон. Это изменение происходит в устройстве или в окислительно-восстановительном процессе хромофора и выражается в изменении устройства белка или его деятельности, и вызывает каскад трансдукции сигнала. Примеры для пигментов фоторецепторов, находящиеся в сетчатке глаза: например, родопсин (версия Миг) (относящийся к сложным белкам хромопротеинам) вызывает жёлтый цвет; например, криптохром (cryptochrome), и билин (bilin) вызывает голубой цвет растений. Относительно фотопигмента цианолаба, который ещё не выделен учёными, то в результате фототрансдукции сигнала, пигмент, который светится в жировых капельках синим цветом, увиден и его можно назвать любым именем, важен цвет, который виден при исследованиях на снимках, и который имеется на синих колбочках-S. Не исключено, что он близок к меланопсину, билину. В любом случае, он является разновидностью фотопигмента опсина (См. Белок и передача зрительных биосигналов (версия Миг)) [цитата, необходимая] Фиг. К; Фигура «синей» колбочки сетчатки примата при электронной микроскопии и её связи с ячейками ганглиозного слоя сетчатки.[4]
Фиг. К; Фигура «синей» колбочки сетчатки примата при электронной микроскопии и её связи с ячейками ганглиозного слоя сетчатки.[4]


Например, проведенное исследование модели протеина (2009) «bestropin-1» — модель белка bestropin-1 в мембране рецепторов (приспособленный от Milenkovik), но только в четырёх областях, расположенных в мембране колбочек сетчатки человека, что говорит о процессах мутации и клетки белка, что показано в цвете и обозначено в цвете. (См.рис. П).[7]
- По вопросу хроматической аберрации, визуального восприятия предметных точек, остроты зрения, о строении фокальной поверхности сетчатки (мозаика, блоки фоторецепторов) и др.
При учёте хроматических аберраций глаза учитывается высота мембраны колбочки в 50мкм. Любой сфокусированный монохроматический луч спектра может сфокусироваться на разной высоте мембраны, даже если все колбочки строго одинаково расположены в фокальной поверхности сетчатки. Однако и в этом случае мембраны колбочек на изображении среза сетчатки расположены не на одной высоте. Палочки расположены расположены выше колбочек. Подробнее смотри выше раздел «Мозаика экстерорецепторов сетчатки». При этом вопрос аберраций света в глазе имеет фундаментальный характер цветного зрения. Так как при оппонентном выделении колбочками основных световых сигналов S,M,L КЗС в блоке, например, трёх колбочек, а сфокусированный луч предметной точки фокусируется в виде кружка нерезкости равной или больше кружка нерезкости (3‒5мкм), то в режиме оппонентного отбора наиболее ярких монолучей, каждая колбочка выделяет один из трёх основных лучей S,M,L, которые посылаются в мозг. При этом каждый сфокусированный луч ложится в конусной мембране в сечениях в зависимости от его длины волны с размером диаметра поперечного сечения фронта волны. Например, синий луч проходит до наименьшего размера сечения конуса конусной мембраны колбочки, то есть у вершины конуса. Или в пределах всей длины мембраны в 50мкм лучи фокусируются точно на разных высотах, что исключает погрешности хроматической аберрации и обеспечивает глазу выдавать биосигналы лучей предметных точек в мозг без искажений. Откуда оптическая система глаза не требует от преломляющей системы хрусталика и зрачка устройства по типу современных анастигматов как в фотографии. Где все лучи фокусируются в одной фокальной плоскости.
Дополнительно цветное зрение у людей и животных связано с вопросами саморегуляции организма, с наличием совокупного движения, поведения биологических систем, которые должны быть замкнутыми и одновременно должны иметь модулятор движения, связывающий организм и его внутренние системы с подвижностью, колебаниями внешней среды, из которой поступает сигналы, возбуждающие по амплитуде или частоте, требующие взаимодействия с элементами нервных узлов. Это относится также и к сетчатке с фоторецепторами. Живая ткань не может быть статичной и колбочки также подвержены ретиномоторной реакции, что сказывается на восприятие, на зрительный процесс в режиме дневного и сумеречного освещения (См. Ретиномоторная реакция фоторецепторов (версия Миг)).
На основании флюоресцентной наноскопии срезов живой сетчатки доказано, что восприятие света и цвета основано на работе фоторецепторов сетчатки, состоящей из мозаики блоков («клеток») с разным количеством колбочек, которые воспринимают основные монохроматические лучи спектра света предметной точки индивидуально в зависимости от строения сетчатки. Например, у птиц мозаика сетчатки состоит из блоков, содержащих систему колбочек «четырехроматик» (четыре колбочки), у человека — "трихроматизм" (три колбочки). (см. рис.3а,6). То есть в каждом блоке мозаики сетчатки могут содержаться по три, четыре, пять и т. д. колбочек, воспринимающих и выделяющих оппонентно основные лучи S,M,L (RGB).
- Также на основании опубликованных данных исследований профессора департамента офтальмологии функциональной нейроанатомии сетчатки R.E.Marcа (2009), то уже в 1966 году при электронной микроскопии сетчатки получены данные «синих» колбочек приматов (см. фиг. К).[4](См.Труды доктора Р.Е.Марка и его лаборатории).
Например, из условия наличия 6 млн колбочек в жёлтом пятне (у человека), на площади 6 мм², которые воспринимают цвет, можно на базе известных данных показать, что одна колбочка не в состоянии выдать нужную информацию цветов, сфокусированной на сетчатку предметной точки. Известно, что разрешающая способность нормального глаза при чтении с расстояния 250 мм находится в пределах 0,072‒0,200 мм и в зависимости от освещённости и индивидуума примем среднестатистическую величину оценок разрешающей способности оптических приборов, среднестатистических глупп взрослых людей, проходящих тестирование (водителей транспортных средств, военнослужащих и т. д.) с показателем 0,0896мм (При остроте зрения 0,8). Основное количество фоторецепторов желтого пятна в центре сетчатки (~6млн) имеет площадь ~ 5,6‒6 мм², (то есть оптическое изображение содержит 1000000 (1мгп) разных цветовых точек), где: расстояние между одноименными точками (фоторецепторами или «пикселами») равно примерно 3‒4мкм (диаметральный наибольший размер основания конуса мембраны колбочки). Рассматриваемые визуально, например, две предметные точки в виде облачков, кружков нерезкости с просветом, равный диаметру кружка нерезкости, которые глаз чётко видит. (Более подробно см. Современный взгляд на световосприятие и цветное зрение (версия Миг), Острота зрения (версия Миг)).


Из условия остроты зрения глаза — способности рассмотреть две точки с просветом между ними на определённом расстоянии (например, при остроте зрения человека 1,0 оно равно 72,5мкм), одна колбочка этого сделать не может. Откуда всегда рассматривается две точки с просветом минимум двумя колбочками или палочками, которые образуют блоки, например, RGB мозаики сетчатки (См. Цветное зрение у птиц (версия Миг)). Как показано выше, диспергированные монолучи предметной точки с диаметром кружка нерезкости примерно 7мкм накрывают три колбочки минимум одного блока с диаметром 6,9мкм (где диаметр мембраны колбочек примерно = 3‒4мкм). Таким образом, в любом случае предметные точки участвуют в восприятии изображения блоком из трёх колбочек мозаики сетчатки. А это значит, что 6 млн колбочек жёлтого пятна объединены в 1,5‒2 млн блоков RGB, которые оппонентно трансдукцируют выходные биосигналы красного, зелёного, синего цвета под управлением головного мозга и фоторецепторов ipRGC, попадающие в зрительные отделы головного мозга через жировые капельки и далее по нервным путям. (Около 1 млн стволов). При этом часть сигналов из 1,5‒2 млн фильтруется, подавляется в блоках при трансдукции RGB под управлением головного мозга и фоторецепторов ipRGC. Принято, что все колбочки одинаковые, каждая колбочка работает в оппонентном отборе основных лучей спектра S,M,L (RGB) предметной точки, которые на неё сфокусированы. При этом наиболее ярких лучей.
Ссылкой на источник в виде:
- Единственные результаты, так ни кем и не оспоренные на сегодняшний день, были получены группой учёных W. B. Marks, W. H. Dobelle, E. F. Mak Nichol. Их исследование подробно описано в работе «Visual Pigments of Single Primate Cones» W. B. Marks, W. H. Dobelle, E. F. Mak Nichol Department of Biophysics, Johns Hopkins University, Baltimore, Maryland 21218. Science, Vol. 143, p. 1182, 1964., и что при освещении только её узкополосным излучением (вырезанным из широкого непрерывного спектра), по спектру отражения можно определить именно ту область, которую поглощает колбочка (область чувствительности колбочки). Для этого использовался широкополосный спектр оптического излучения из которого монохроматором вырезался очень узкий участок спектра. Далее этот монохромный спектр фокусировался в точку размером меньше, чем геометрический размер самой колбочки. Эти точки сканировали в сетчатке живого глаза.
Как видим, источник, где проведенное исследование в 1964 году при сканировании сетчатки не учитывалась работа мозга и фоторецептора ipRGC (открытые в 2007 году) с фотопигментом меланопсином, которые дозируют, фильтруют область сильных фотонов коротковолновых монохроматических лучей (фиолетовых-синих) не случайно не позволили в то время обнаружить колбочек, трансдукцирующих синие лучи света.
- По вопросу дефектов цветовосприятия.
Именно, с открытием третьего фоторецептора сетчатки ipRGC (2007) учёные получили возможность глубже вникнуть в природу отклонений в восприятии цвета зрительной системой. Не рассматривая в комплексе работу колбочек, мозга, фоторецепторов ipRGC, конечно, не возможно оценить цветное зрение. Только с позиций трихроматизма — теории трёхкомпонентного цветного зрения практически решаются все вопросы цветовосприятия.
Таблица видов дальтонизма[править | править код]
Эта таблица для обсуждения взята из англо-вики, с сокращениями и частичным переводом. После окончания работы над ней, она может быть введена в статью Дальтонизм. Полезно добавлять в неё более новые научные источники с описанием, новые факты (в том числе о распространённости отдельных видов дальтонизма в различных популяциях), чтобы эта табл. действительно стала важной частью статьи о дальтонизме.
| Мужчины | Женщины | всего | Ссылки | |
|---|---|---|---|---|
| 1. Red-green (в общем) | 7 to 10 % | — | — | [8][9] |
| 1.1 Red-green (кавказцы) | 8 % | — | — | [10] |
| 1.2 Red-green (азиаты) | 5 % | — | — | [10] |
| 1.3 Red-green (африканцы) | 4 % | — | — | [10] |
| 2. Monochromacy | — | — | — | |
| 2.0 Rod monochromacy (dysfunctional, abnormally shaped or no cones) | 0.00001 % | 0.00001 % | — | [11] |
| 3. Dichromacy | 2.4 % | 0.03 % | 1.30 % | [8][11] |
| 3.1 Protanopia (red deficient: L cone absent) | 1 % to 1,3 % | 0.02 % | — | [8][11] |
| 3.2 Deuteranopia (green deficient: M cone absent) | 1 % to 1,2 % | 0.01 % | — | [8][11] |
| 3.3 Tritanopia (blue deficient: S cone absent) | 0.001 % | 0.03 % | — | [11] |
| 3. Неполный трихроматизм | 6.3 % | 0.37 % | — | [11] |
| 3.1 Protanomaly (red deficient: L cone defect) | 1.3 % | 0.02 % | — | [11] |
| 3.2 Deuteranomaly (green deficient: M cone defect) | 5.0 % | 0.35 % | — | [11] |
| 3.3 Tritanomaly (blue deficient: S cone defect) | 0.01 % | 0.01 % | — | [11] |
Из таблицы ясно, отчего некоторые виды дальтонизма, возможные теоретически, не выявлены в клинических исследованиях — их частота пропорциональна произведению вероятностей «выпадения» отдельных типов рецепторов (вероятность одновременного наличия дефективных генов). Alexandrov 14:32, 16 мая 2011 (UTC)--Миг 07:58, 19 мая 2011 (UTC)
См.также[править | править код]
- Теория трёхкомпонентного цветного зрения (версия Миг)
- Линейное уравнение цветного зрения и цветового пространства
- Нелинейная теория зрения
Примечания[править | править код]
- ↑ http://humbio.ru/humbio/ssb/0008994b.htm#001181d1.htm
- ↑ http://humbio.ru/humbio/ssb/001181d1.htm
- ↑ http://eps.dvo.ru/bm/2006/1/pdf/bm-055-059.pdf
- ↑ а б в г д "Seeing in color". Prometheus. Дата обращения: 8 сентября, 2012.
{{cite web}}: Проверьте значение даты:|accessdate=(справка)К:Википедия:Ошибки CS1 (даты) - ↑ Marc RE, Sperling HG. (22 апреля, 1977). "Chromatic organization of primate cones". Дата обращения: 8 сентября, 2012.
{{cite web}}: Проверьте значение даты:|accessdate=and|date=(справка)К:Википедия:Ошибки CS1 (даты) - ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://dare.ubn.kun.nl/bitstream/2066/74414/1/74414.pdf
- ↑ а б в г "Prevalence and Incidence of Color blindness". WrongDiagnosis.com. 22 июня 2008. Дата обращения: 10 июля 2008.
- ↑ "Color Blindness: More Prevalent Among Males". Hhmi.org. Дата обращения: 16 апреля 2009.
- ↑ а б в Masataka Okabe, Kei Ito (15 февраля 2008). "Colorblind Barrier Free". J*Fly. Дата обращения: 10 июля 2008.
- ↑ а б в г д е ё ж з Archive.org
| Это незавершённая статья. Вы можете помочь проекту, исправив и дополнив её. |




