Теория трёхкомпонентного цветного зрения (авторская основного пространства участника Миг)





Теория трёхкомпонентного цветного зрения основана на базе физики цвета (RGB) и взаимодействия с ним живой среды обитания — биологической, а также на основе колориметрических, гистологических, генетических исследований[5], на основе достижений в этой области за последние 30-35 лет, на основе биофизики, биохимии, пересмотра основ прежнего сложившегося процесса зрения с разных точек зрения:
- С точки зрения биологической, в области цветного зрения начиная с 1966 по 2009 годы (Лаборатория Р.Е.Марка) с основным экспериментальными данными исследований живой клетки, на срезах сетчатки установлена работа колбочек и (S,M,L) в блоке RGB и палочек. В условиях дневного освещения (цветного зрения) — колбочек и работа палочек, в период сумеречного и ночного освещения (не цветного) изолировано от колбочек. Работа фоторецепторов связана с полученными данными работы видоизменяющихся разновидностей фотопигментов на базе белков опсинов:
- Кон-опсины(конопсин) — у колбочек,
- Род-опсин(родопсин) — у палочек.
- С точки зрения чисто физической на базе труда (2011 года) учёного физика[6] Джеральда К. Хата, который на базе физики взаимодействия света с внешними долями мембран фоторецепторов сетчатки глаза, где в основном осуществляется первичное взаимодействие со светом на наноуровне («nanostructural») (рецепторное) и доктора физика Джона Медейроса[4], который рассмотрел работу внешних долей мембраны колбочек и палочек с точки зрения аналогичной работе волноводов конической и цилиндрической формы в среде прозрачного тела глаза (жидкая среда).
- В конечном итоге, физики Дж. К. Хат и Джон Мейдерос пришли к общепринятому принципу трихроматизма. (См. Пересмотр традиционных взглядов на зрительный процесс, Работа мембраны колбочек и палочек как волновод).
Так с точки зрения чисто физичекой физика, доктора К.Хата работа сетчатки укладыается в направление, которое подводит к «nanostructural», когда свет рассматривается с вопросом как электромагнитная волна в классической физике, который проходит через пространственную размерность (названной «нано-антеннами») и фильтруется в виде трёх основных цветов RGB в мембранах колбочек и палочек — образуя систему трихроматизма.
Падающий на сетчатку свет и его восприятие следует подразумевать, что при поглощении света имеет место взимоотношение в пространственной размерности элементов между смежной колбочкой и палочкой, но не в виде чистого кванта и подразумевается, что фотоны взаимодействуют непосредственно в пределах образования — фоторецепторов: 1 колбочка:8 или 6 палочки вокруг неё!.
- Последние данные Джеральда К. Хата (2010-2011) не противоречат полученным результатам известных учёных всей предыдущей истории цветного зрения на базе биофизики, а только их уточняет, исправляет во взаимодействии с ним.
- И с точки зрения доктора, физика Джона Мейдероса, внешние доли мембран колбочек и палочек он рассматривает как биологические волноводы, которые воспринимают световые лучи с фиксацией их в сечениях волноводов, равных размерам поперечного сечения фронта световой волны в обратном порядке прохождения их (внешних долей мембран колбочек) в жидкой среде, нежели в обычных оптических волноводах в воздухе. (Связано с влиянием показателей преломления сред).
Введение[править | править код]
За последние (примерно 30-35 лет) на основе биофизики, биохимии пересмотрены основы прежнего сложившегося процесса зрения с разных точек зрения:
- С точки зрения биологической, в области цветного зрения начиная с 1966 по 2009 годы (Лаборатория Р.Е.Марка) с основными экспериментальными данными исследований живой клетки, на срезах сетчатки установлена работа колбочек (S,M,L) в блоке RGB и палочек. Установлено, что в условиях дневного освещения (цветного зрения) работают колбочки и в период сумеречного и ночного освещения (не цветного) работают палочки, в режиме изолированном от колбочек. Работа фоторецепторов связана с видоизменяющимися, разновидностями фотопигментов на базе белков опсинов (См. Ретиномоторная реакция фоторецепторов):
- Кон-опсины (конопсин) — у колбочек,
- Род-опсин (родопсин) — у палочек.
- С точки зрения чисто физической на базе труда (2011 года) учёного физика[7] Джеральда К. Хата предложено рассматривать взаимодействия света с внешними долями мембран фоторецепторов сетчатки глаза на основе первичного взаимодействия со светом (рецепторное) на уровне «нано-антен». По данным доктора физика Джона Медейроса[4] рассмотрена работа внешних долей мембраны колбочек и палочек аналогично работе волноводов конической и цилиндрической формы в среде прозрачного тела глаза (жидкая среда).
- В итоге, физики К.Хат и Джон Мейдерос пришли к общепринятому принципу трихроматизма. (См. Пересмотр традиционных взглядов на зрительный процесс, Работа мембраны колбочек и палочек как волновод).
Например, важными открытиями являются: работа фоторецепторов колбочек, палочек — Ретиномоторная реакция фоторецепторов, флюоресцентная микроскопия сечений живой сетчатки цыплёнка (Цветное зрение у птиц), открытие третьего ганглиозного фоторецептора сетчатки ipRGC c фотопигментом меланопсином (Фоточувствительные клетки сетчатки ipRGC), отчёт лаборатории Р.Е.Марка в области цветного зрения начиная с 1966 года (Лаборатория Р.Е.Марка) с основным экспериментальными данными в области трихроматизма, работой колбочек (S,M,L) в блоке RGB в условиях дневного освещения (цветного зрения) и палочек, в области сумеречного зрения (не цветного).
Пока выделены у колбочек разновидности фотопигмента колбочек йодопсина — фотопигменты хлоролаб и эритролаб, у палочек — родопсин.
Фотопигмент с общим названием Йодопсин содержащится в колбочках и участвует в механизме цветового зрения. Согласно теории цветного зрения трёхкомпонентной в настоящее время принято, что существуют три вида этого пигмента и сетчатка глаза соответственно содержит одинаковые колбочки, которые в зависимости от воздействия световых лучей с разными длинами волн (S,M,L) трансдукцируют биосигналы в мозг оппонентно отобранных основных лучей спектров света предметных точек: голубого, зелёного и красного цвета. Однако пока известно, что йодопсин находится во всех колбочках глаза. Он включает в себя фотопигменты хлоролаб, который поглощает лучи жёлто-зелёной части спектра) и эритролаб который поглощает лучи жёлто-красной части спектра)). На сегодня пока не выделены разные все типы фотопигментов. Обнаружить присутствие третьего только теоретически предсказанного синечувствительного фотопигмента «цианолаба», который (по трёхкомпонентной теории цветового зрения) содержится в колбочке «в третьем случае приёма синих лучей», пока не удалось (ни у дихроматов, ни у трихроматов), что объясняется жёлтой пигментацией в районе центральной ямки, которая по мнению исследователей затрудняет идентифицировать фотопигмент при измерении, отборе, фототранслукции сигнала при ввоздействии на мембрану колбочек-S лучей фиолетово-синего спектра.
Однако установлено, что в восприятии отделных цветов (S,M,L) участвуют фоторецепторы с разновидностями фотопигментов на базе опсинов:
- S-спектр — фотопигменты β кон-опсин, меланопсин (фоторецепторы: Колбочки-S, ганглиозные клетки ipRGC.) (Фотопигмент восприятия синих лучей S-спектра колбочками-S пока не выделен, т.к. энергонасыщенные синие лучи при взаимодействии с фотопигментом фоторецепторов колбочек-S c повышеной чувствительностью и скоротечностью процеса фототрансдукции на фоне жёлтого цвета ямки представляют затруднения в индентификации разновидности фотопигмента β кон-опсина[3]),
- M-спектр — фотопигменты γ кон-опсин с разновидностью йодопсина (фоторецепторы: Колбочки-M) — хлоролаба, чувствительного к жёлто-зелёным частям спектра,
- L-спектр — фотопигменты ρ кон-опсин разновидность йодопсина (фоторецепторы: Колбочки-L) — эритролаба, чувствительного к жёлто-красной частям спектра).
- Также на основе проведённых фундаментальных исследований академика биохимика М.А.Островского в области работы фотопигмента родопсина в палочках[8] (Фотобиологический парадокс зрения) можно утверждать, что в настоящее время уже не стоит вопрос о принципах трёхкомпонентной теории цветного зрения в целом. Современные лаборатории в области зрительной системы занимаются в настоящее время вопросами работы всех нервных клеток глаза и мозга, участвующие в создании, проведения биосигнала предметной точки оптического изображения на рецепторном и нейронном уровнях до попопадания в мозг. Т.е. ведётся работа по созданию атласов прохождения сигналов с целью возможности диагностирования, лечения заболеваний зрительной системы. (См. Лаборатория Р.Е.Марка).
Что касается других вопросов, это:
- Создание и совершенствования теорий цветного зрения, которые в общем изложены в статье Теории цветного зрения, и частично — в Истории оптики.
- То, что мы ощущаем как цвет, представляет собой комбинированное воздействие:
- 1) Спектрального распределения светового потока из дающего энергию источника света;
- 2) Физических и/или химических свойств всех материалов, пропускающих или отражающих световой поток (по меньшей мере часть светового потока, переориентированную в сторону глаза);
- 3) Физиологической реакции глаза на световой поток, включающей в себя нервные импульсы, передаваемые в ту часть коры головного мозга, которая отвечает за зрение;
- 4) Переработки нашим мозгом этих сигналов в сочетании с сигналами из соседних областей поля зрения, нашими воспоминаниями о сходных ситуациях, имевших место в прошлом опыте[9].
- Хотя это достаточно общее высказывание в целом верно, но для того, чтобы «теория цвета» стала продуктивной, требуется дальше развить и углубить, связать с фактами каждую модель уровней цветовосприятия.
Требования к теории восприятия света и цвета[править | править код]
Факты и их объяснение[править | править код]
К главным вопросам, раскрывающих суть трёхкомпонентной теории цветного зрения, относятся:
- Экспериментальные гистологические научные данные, проводимые при микроскопии на базе современных флюоресцентных наноскопов с использованием живых клеток (сечений сетчатки глаза) с разрешающей способностью 1-10нм, в цветовом трёхмерном пространстве на атомно-молекулярном уровне. (См. Цветное зрение у птиц);
- Колориметрия;
- Цветоведение;
- Оптика;
- Физиология и анатомия человека;
- Биохимия восприятия света;
- Генетика;
- Психология восприятия цвета.
Все указанные факторы должны быть непротиворечиво объединены с помощью наиболее понятного понятийного и математического аппарата.
Среди фактов, которые должны быть учтены:
- Данные последних исследований строения сетчатки при микроскопии живых клеток (рентгеноскопии, флюоресцентной наноскопии);
- Учёт спектра чисто земных изображений, который может быть в виде двух основных компонентов:
- Цветной контраст между короткими и средними длинами длин волн спектра;
- Цветной контраст между средними и длинными частями длины волны спектра.
- Должна быть использована информация о приматах и человеке, и чтобы извлечь и использовать эту информацию, требуется:
- Определение в видимой части соответствующей области длины волны;
- Учёт морфологии (схемы) сетчатки глаза, которая должна приспосабливаться, воспринять спектральную информацию и быть эффективно переданной в мозг.
Животные, приматы, зрительные системы которых отвечают этим требованиям, наделены способностью цветного зрения[10][11].
- Должно быть учтено и выдвинуто на первый план недавнее продвижение понимания трех аспектов видения цвета trichromaticу у приматов (2002г):
- 1) поведенческое значение trichromacy,
- 2) развитие и генетика фотопигментов,
- 3) относящиеся к сетчатке глаза кругообороты, которые создают красно-зеленые и синие-желтые цветные каналы в зрительном нерве[12].
Общие требования[править | править код]
Теория цветного зрения трёхкомпонентная как и любая теория может считаться полезной только при выполнении следующих условий:
- теория должна строиться на базе объективных, подтверждённых экспериментальных данных,
- на основе использования информации современной микроскопии и гистологического анализа (например, флюоресцентной микроскопии живых тканей) на атомно-молекулярном уровне, в цветовом 3D пространстве,
- теория должна основываться на физических законах,
- научные способы (включая Научное моделирование) должны содержать совокупность основных способов и методов решения задач в рамках получения новых знаний,
- умозаключения и выводы должны получаться с помощью правил и принципов рассуждений на основе эмпирических (наблюдаемых и измеряемых) данных об объекте[13]
- учёт работы всех фоторецепторов сетчатки и фотопигментов, их нейронные связи между собой и мозгом,
- объяснение основных и дополнительных цветов, понимание их влияния на контрастность, яркость изображения,
- законов смешивания цветов (например, аддитивный синтез цвета, субтрактивный синтез цвета),
- основываться на биофизических и биохимические законах,
- описываться математическими зависимостями в трёхмерном 3D пространстве.
Феноменология цветовосприятия[править | править код]


Различные феномены цветного зрения показывают, что зрительное восприятие света, цвета зависит не только от вида воздействующих источников света и цвета и работы фоторецепторов, но также от способа переработки световых сигналов в зрительной системе. Различные участки видимого спектра света кажутся нам по-разному окрашенными, при этом отмечается непрерывное изменение ощущений восприятия цвета при переходе от фиолетового и синего через зелёный и жёлтый цвета к красному. Одновременно человек может воспринимать цвета, которые не спектральные (отсутствующие в спектре), например, пурпурный цвет, получаемый при смешении красного и синего цветов. Различные физические способы цветового воздействия зрительной системы могут приводить к идентичному восприятию цвета. Например, монохроматический жёлтый цвет не отличается от соответствующей смеси монохроматических зелёного и красного цветов. Или при воздействии на зрительную систему нужным набором цветов RGB («белого» цвета, например, солнечного) на диске, то при его вращении мы увидим диск белого цвета, и выполнив тоже самое, но c набором XYZ — мы увидим чёрный цвет (точнее тёмно-коричневый) (см. рис.3).
Феноменологию цветовосприятия описывают законы цветного зрения, выведенные по результатам психофизических экспериментов. На основе этих законов за период более 200 лет было разработано несколько теорий цветного зрения. Однако приблизительно в последние 35 лет появилась возможность непосредственно проверить эти теории методами электрофизиологии путем регистрации биоэлектрической активности одиночных фоторецепторов зрительной системы, а также расширить область взаимодействия и количество фоторецепторов сетчатки и мозга в цветовосприятии. Например, в 2007 году признано открытие 1991 года экстерорецепторов ipRGC (не колбочки и не палочки, расположенных в ганглиозном слое на пути световых лучей оптического изображения в сетчатке глаза).

.
В связи с проведенными ранее исследованиями сетчатки при помощи рентгеноскопии (у приматов)[16] и проводимые в настоящее время исследования живых клеток и в том числе клеток сетчатки (колбочек, палочек и ipRGC) на базе нового микроскопа — Флюоресцентного наноскопа с разрешающей способностью 1-10 нм, на атомно-молекулярном уровне, появилась возможность получать новые данные исследований в цвете (это важно для оценки восприятия цвета экстерорецепторами, чувствительными к определённой длине волны спектральных зон S,M,L) с данными стереоизображений на мониторе в заданном масштабе, а также новых методов измерения цвета (см. Измерение цвета) (см. рис. 4а). Так уже проведены работы при исследовании цветного зрения у птиц на живых клетках сечений сетчаток (см. Цветное зрение у птиц) в 2006—2008 годах[5]. Открытие третьего типа экстерорецепторов ipRGC в сетчатке дало возможность более глубже взглянуть в тайны работы зрительной системы, уточнить детали процесса восприятия света и цвета. Также удалось связать работу экстерорецепторов сетчатки (палочек, колбочек, ipRGC), расположенных в разных слоях, функционирующих в биологической среде дифференцировано и в единой системе с участием коры головного мозга (осознанно).
В данной биологической системе функционирует прямая и обратная связь между экстрорецепторами и мозгом, при которой образование оптического изображения происходит через несколько этапов: создание, получения «первичного» оптического изображения в сетчатке (на фоторецепторном уровне — палочек, колбочек) с передачей сигналов от изображения предметных точек в головной мозг. Нейронные сети, в режиме принципа «оппонентности» (см. Оппонентная теория цветного зрения) формируют уже в нашем сознании оптический образ — «изображение» (в зрительных отделах головного мозга). При этом оптические изображения формируются перекрёстно в правом и левом полушарии головного мозга соответственно от левого и правого глаза в стерео, т.е. бинокулярном зрении. И в основе трёхкомпонентного принципа RGB (трихроматизм) заложено:
- 1) Создание чёрно-белых оптических изображений в сетчатке «контурно» — (рецепторный уровень),
- 2) Создания цветных оптических изображений в мозгу — (нейронный уровень).
Процесс возникновения ощущения цвета можно кратко представить следующим образом. Согласно традиционному подходу, свет представляет собой сложный набор предпосылок для восприятия цветных лучей. Пока свет от источника или отражающей поверхности не достиг рецепторов цветного зрения сетчатки (колбочки), считается, что цвета нет. Колбочки избирательно чувствительны к синей, зелёной и жёлто-красной частям спектра. Кроме этого существует палочковая система фоторецепторов, реагирующая на слабую освещенность и обеспечивающая сумеречное и ночное зрение.
Возникновение цветовых ощущений как механизм цветоразличения хорошо описывается в известной трёхкомпонентной теории Юнга-Гельмгольца. Последняя объясняет необходимость и достаточность классической триады основных цветов RGB (красного, зелёного и синего) для получения цветов видимого спектра путём аддитивного смешения (Педхем, Сондерс, 1978). Например, на этом основана технология получения цвета в кинескопе. Трёхкомпонентная теория оказалась полезной и в качестве основы для различных процессов воспроизведения цвета и была развита с помощью законов Гроссмана в метод колориметрии МКО (CIE — в английской транскрипции). От первичных детекторов (рецепторный уровень) сетчатки возбуждение передаётся далее на группу градуальных нейронов, составляющих второй детекторный уровень (нейронный уровень — мозг)(Соколов, Измайлов Ч. А., 1984; Измайлов, Соколов, Черноризов, 1989). В настоящее время считается, что существует три вида детекторов этого уровня: красно-зелёный, сине-жёлтый и чёрно-белый (яркостной) (Педхем, Сондерс, 1978), хотя существует обоснованное мнение, что их должно быть не менее четырех (Соколов, Измайлов Ч. А., 1984; Измайлов, Соколов, Черноризов, 1989). Это связано с выявлением в структуре процесса цветоразличения не только в виде оппонентного отбора цвета яркого сигнала — анализ по яркости, но и так называемого униполярного темнового механизма, то есть анализатора белизны, что соответствует ощущению насыщенности цветового тона. Как бы то ни было, на этом уровне характер обработки цветового раздражения хорошо укладывается в теорию оппонентности Эвальда Геринга. Эта теория основывается на существовании уже не трёх, а четырёх основных цветов: красного, зелёного, жёлтого и синего, остающихся неизменными по цветовому тону при различных стимульных условиях и субъективно выделяемых большинством людей в качестве главных элементов цветовой гаммы. На основании первой части теории Геринга была разработана современная Натуральная цветовая система (NCS) (Тонквист, 1993)[17].
Дальнейшая обработка информации в цветовом анализаторе предполагает процесс сличения раздражителя с узкополосным эталоном (см. обзор: Демидов, 1987), позволяющий идентифицировать мелкие цветовые поля на фоне крупных. Существует также гипотеза о дублировании всего множества селективных детекторов цвета нейронами образной памяти (Соколов, Вартанов, 1987). Пройдя весь сложный путь от глаза до зон цветового анализатора в коре, трандукцированные сигналы, воспринятые рецепторами глаза, превращаются в то, что мы воспринимаем и называем как Цвет. Этот путь в настоящее время может быть представлен как последовательная сортировка количественных данных (частот спектра) на некие всё более дробные качества в форме специфических реакций полей детекторов или ансамблей нейронов. Таким образом, данные классических и современных исследований позволяют мыслить Цвет не только в форме электромагнитных колебаний, но и в терминах спайковой (то есть в конечном итоге тоже электромагнитной) активности строго определенных нейронных структур мозга. До сего дня не решена проблема преодоления границы между физическими закономерностями, организующими работу физического аппарата восприятия, и возникновением психического феномена: «„Ни теория Юнга-Гельмгольца, Теория Э.Геринга не могут полностью объяснить, как сигналы преобразуются в мысленный образ объекта“ (Тонквист, 1983)»[18].
- Данный подход глубоко и безнадёжно ущербеный, так как он погряз в так называемой психофизиологической проблеме. Суть которой в том, что при данном способе рассуждений как раз имеет место непроходимая стена между ощущением и образом. За психо-физиологической проблемой стоит проблема психо-физическая. Психо-физическая проблема — камень преткновения всех концепций, изолированно изучающих душу и тело, человека и мир. Это — тупик из тупиков любого негумманистического подхода к Человеку и Природе[19].
Тем не менее, с точки зрения материалистической в настоящее время на первый план выходит недавнее продвижение понимания трёх аспектов видения цвета трихроматизма (trichromacy), которая принята на базе данных исследований у приматов (2002 г):
- 1) поведенческое значение трихроматизма,
- 2) развитие и генетика фотопигментов,
- 3) относящиеся к сетчатке глаза кругообороты, которые создают красно-зелёные и сине-жёлтые цветные каналы в зрительном нерве (см. Участник:Миг/Механизмы цветовосприятия у приматов, Лаборатория Р.Е.Марка)[20].
Принцип трихроматизма — основа цветного зрения[править | править код]





Трёхкомпонентная теория цветовосприятия исторически основана на многие гипотетические теории, предсказательная сила которых была невелика. Даже гениальные предположения учёных прошлого были в основном умозрительны, так как естественные науки прошлого не могли расширяться, т.к. не обладали в своё время необходимым инструментарием; не был сформирован понятийный аппарат, невозможно было провести многие критически важные эксперименты в силу недостаточной разрешающей способности оптико-механических приборов (например, микроскопов). Бурное развитие естественных наук можно отнести начиная с ХIХ столетия, когда специалисты в области биологии, химии и физики сделали огромный прорыв в естественных науках. (См.Участник:Alexandr/Современные концепции световосприятия и цветного зрения).

Цвета в этом файле определяются, используя RGB. Области вне треугольника не могут быть точно ограничены, потому что они — вне гаммы RGB, поэтому интерпретировались. Изображённые цвета зависят от цветного места устройства, на котором Вы рассматриваете изображение, и поэтому здесь не можете быть строго, точного представления цвета в специфическом положении. Различные участки спектра, формируют выходные биоэлектрические сигналы в каждом из (см. рис.2а,3а) трёх типов фоторецепторов в различной степени. Жёлтоватый цвет, например, стимулирует L и М колбочки одинаково, а синеватый — Булочками-колбочками-S. Красный цвет, с другой стороны, стимулирует колбочки L намного больше, чем М., а колбочки S — зависят и возможно стимудируются фоторецепторами ipRGC при поддержке мозга в режиме фильтрации ими сильных УФ лучей.; синий-зелёный цвет стимулирует L колбочки слабее, чем колбочки M и S, а вот колбочки M и S — сильно. На нейронном уровне (мозг) контурные сигналы (аддитивно синтезированные), поступающие из сетчатки (рецепторного уровня), трансдукцируют в цветную.
На базе полученных новых данных максимумов спектральной чувствительности фотопигментов клеток сетчатки (S,M.,L), палитра цветов, которую может различать глаз, работающий в соответствии с трёхкомпонентной теорией зрения с учётом всех фоторецепторов сетчатки глаза и мозга, имеет вид согласно диаграммы палитры. На рис.2а,3а,Fig.15 видим области цветовой палитры, характеристику графиков кривых цветного восприятия колбочками, которую анализируют или синтезируют при известных максимумах чувствительности пигментов рис.3, Fig. 15 (колбочек) в соответствии с оппонентным отбором основных цветов по теории цветного зрения трёхкомпонентной.
Наличие фотопигментов L и М колбочек, где L и М закодировано на хромосоме X; дефектное зашифровывание их приводит к двум самым общим формам дальтонизма. Ген OPN1LW, который закодировал для пигмента, и который отвечает на желтоватый цвет, является очень полиморфным (недавнее исследование Verrelli), Tishkoff нашел 85 вариантов при исследовании 236 мужчин {полиморфный}[23]. Десять процентов женщин[24] имеют дополнительный тип гена, и таким образом возможно степень различия цвета у них может быть (а может и не быть) несколько иной[25].
Клетки ipRGC, расположенные в сечатке, в зоне Ganglioniс layer и рефлекторно связанны с палочками и колбочками, имеют прямую и обратную сязь с мозгом. Они учавствуют в обработке информации поступающей изначально при зрении, являясь фильтром УФ падающих лучей и взаимодействуют с фоторецепторами сетчатки (колбочки и палочки) и мозгом, т.к. они непосредственно своими синапсами связаны с ними. Содержа фотопигмент меланопсин (воспринимает и фильтрует фиолетово-синие лучи) фоторецепторы ipRGC автоматически участвуют в работе колбочек-S и зрительных отделов головного мозга в создании оптического изображения.
Визуальное цветное зрение[править | править код]
Визуальное цветное зрение — основа современных теорий цветного зрения.
Цветовое зрение происходит в зрительной системе, где инициируется поглощением света с помощью трех различных спектральных классов "шишек" — колбочек. Следовательно, цветовое видение описывается как трёхвариантное восприятие основных цветов или как восприятие, ощущение цвета. Первоначально психофизические исследования показали, что цвета могут быть настроены на использование трех различных систем (праймериз). В 1802 году, Томас Молодых предложил модель, по которой восприятие цвета может быть закодировано на восприятие цвета трёх основные фоторецептоов, но не на кодировании тысяч цветовых рецепторов для отдельных цветов.[26]
Спектральная чувствительность колбочек[править | править код]

Спектральная чувствительность колбочек может быть определена посредством нескольких методов. Два из этих метода включают изолирующую рецепторные (receptoral) ответы (Бейлор et al., 1984) с использованием вычисленных от цвета функции нормалей и дихроматизма или двухкомпонентности восприятия цвета (dichromats) (Смит и покорного, 1975; dichromat является предметом, в основе которого сетчатка имеет одну колбочку с фотопигментом в мембране (photopigment), достаточного для этого), микроспекромитрии (microspectrometry) (Bowmaker и Dartnall, 1980) или на основе отражения световых лучей — денситометрии (Раштон, 1963, 1966). В microspectrometer метод предполагает выделение одной колбочки, пропуская свет через неё. Изменения в передаче различных длин волн может быть использовано для вычисления спектрального поглощения колбочкой или возможность определить изменения в электрическом ответе. Отражение денситометрии включает режиссуру, структуру света в сетчатке и определение изменения в поглощении как функция длины волны. Эти результаты впоследствии используются для расчета спектральных поглощений.
Было принято три класса колбочек в человеческой сетчатке глаза, которые были изолированы от указанных технологий. Эти три класса "шишек" представляют:
- Короткие волны чувствительности S-колбочки (S-конуса),
- Средние волны чувствительности M-колбочки (М-конуса),
- Длинноволновую чувствительность L-колбочки (L-конуса).
У всех разная, но пересекающаяся спектральная чувствительность. Спектральная чувствительность S-колбочек с пиком, примерно, в 440 нм, М-коолбочек — 545 нм и L-колбочек с пиком в 565 нм после исправлений, для предварительной потери света сетчаткой. Хотя различные методы измерения дали результаты в несколько различных вариантах максимального значения чувствительности (рис. 1).

Трёхцветная природа цвета[править | править код]

Трехцветная природа цветового зрения позволяет практически получить любой цвет, при смешении трех основных цветов (RGB). При этом trichromacy цветного зрения является линейной функцией. Это означает, что колориметрические уравнений имеют свойства обыкновенного линейного уравнения.
Цвет, соответствующий количеству Cs стимула S, может быть выражен как:
Cs(S) = C1(1i) + C2(2i) + C3(3i)
где li-3i есть три основных цвета (не обязательно монохроматического цвета спектра) и C1-C3 - трехстимульные значения.
Трехстимульные значения представляют собой количества каждого из трех первичных выборов, необходимых для достижения соответствия для цвета и яркости (рис. 2). Они могут быть выражены в единицах светового потока или потока излучения, или даже произвольный масштаб, при условии, что произвольный масштаба передает относительные пропорции трёх количеств.
Трихроматизм и компонентный характер цветного зрения[править | править код]
- (Данный материал создан совместно с Алекандровым и учтён при написании данной статьи.)
Понятие трихроматизма вводится для цветного зрения, что бы выразить связь причины появления цветного зрения, цвета с биологисеским инструментом, создающий это явление как цветное зрение. В природе все биофизические и биохимические процессы подчинены общим законам, которые мы стремимся раскрыть. Не случайно луч света диспергируется на монолучи (спектр), которые участвуют в зрительном процессе. Однако как оказалось, спектр содержит основные и не основные лучи света. Основными оказались три монолуча: красный, зелёный, синий — RGB или RGB (цветовая модель), т.е.RGB (аббревиатура английских слов Rred, Ggreen, Bblue — красный, зелёный, синий) — аддитивная цветовая модель, как правило описывающая способ синтеза цвета для цветовоспроизведения. Откуда весь набор биологических структур цветного зрения подчинён закону аддитивного синтеза цвета. Например, это три цвета реализуются колбочками. Собственно отсюда и названия трихроматизм, трёхкомпонентный принцип цветного зрения. Это случай совпадения триады лучей и триадой фоторецепторов, участвующих (колбочек) при цветном зрении.
При этом существуют одно, двух, трёх, четырёх и более компонентные структуры цветного зрения, но в любом случае (биологическая) система зрения — глаз в результате эволюции имеет в принципе одинаковые структуры для процесса зрения. Это оптическая система, сетчатка с фоторецепторами и зрительные отделы мозга
На сегодняшний день учёные не гипотетически, а на опытах доказали и получили изображения трёх, четырёх и более типов разных колбочек, воспринимающих основные спектральные монохроматические лучи света. Т.о. такая теория на сегодняшний день является основной. В таблице ЗОНЫ ОСНОВНЫХ ЦВЕТОВ RGB определены спектры поглощения фоторецепторами сетчатки, их спектральные максимумы поглощения видимого излучения света — это S, M.,L, зоны основных цветов трихроматизма (номинальные, наиболее яркие спектральные области работы фоторецепторов сетчатки при цветном зрении).
| Обозначения основных цветов и фоторецепторов | Обозначение фотопигментов (разновидностей опсинов) | Диапазон | Пиковая длина волны |
|---|---|---|---|
| S-Палочки — максимальный фиолетово-синий; | (родопсин) | 450-530 нм | 498 нм |
| S-Колбочки, ганглиозные фоторецепторы ipRGC — синий; | β кон-опсин, меланопсин | 400-485 нм | 437 нм |
| M-Колбочки — зелёный;, желтовато-зелёный; сине-зелёный; (красный-мало); | γ (йодопсин) | 455–610 нм | 533 нм |
| L-Колбочки — красный, желтовато-красный; | ρ разновидность йодопсина | 480–650 нм | 564 нм |
Колбочки и цветное зрение[править | править код]



Ко́лбочки — (англ. cone) клетки фоторецептора в сетчатке глаза являются ответственными за цвет, они функционируют в условиях дневного освещения, в противоположность клеткам фоторецепторов палочек, которые начинают работу с наступлением сумеречного и ночного освещения. (См. Ретиномоторная реакция фоторецепторов). Колбочки плотно упакованы в ямке, но постепенно равномерно редеют к периферии сетчатки[31]
В последнее время (2009-2011) учёные физики, доктора Джеральд К. Хат[32] (см. Центральная ямка сетчатки глаза), John A. Medeiros[33] (см. Работа мембраны колбочек и палочек как волновод) внесли новый подход в интерпретацию цветного зрения с точки зрения физики, которая не противоречит, и даже уточняет некоторые полученные выводы, данные известных учёных, предшественников — биологов, биофизиков, биохимиков.
Сейчас учёные рассматривают колбочку как клетку с внешней долей конусной мембраны, имеющей пространственную структуру, работающей по типу конусного волновода в среде прозрачного тела глаза. Это даёт возможность ей воспринимать сфокусированные лучи предметной точки оптического изображения в очерёдности, как работает конусный волновод в прозрачном теле сетчатки глаза, т.е. в очерёдности (обратной): красные→зелёные→синие→оранжевые и т.д.. При этом идёт отбор основныех спектральных квантованных лучей (S,M.,L,) или RGB, притом не в цвете. Эти отфильтрованные квантованные лучи в виде длин волн предметной точки со своими характеристиками преобразуются, трансформируется со скоростью в фемтосекуду, а затем трансдукцируются в мозг, где сфокусированная квантованная предмтная точка оптического изображения (в квантах энергии её длины волны) не в цвете, сканируется со своими координатами, но уже в цвете. Затем сигналы проводятся по нервным стволам в зрительные отделы головного мозга, где преобразовываются у нас в мозгу в виде аналогового оптического изображения в цвете, стерео изображение. Это бинокулярное оптическое изображение (субъективное), которое создаётся у каждого из нас своё.
Никакой заменитель глаза не способен это оптическое изображение повторить, мы можем с применением современной техники приблизиться к глазу, познать его принципы работы, но в настоящее время существует много непознанных вопросов получения оптического изображения на сетчатке, в мозгу. Самое важное, созданная техника не может работать как пространственный биологический фоторецептор — конусная внешняя доля мембраны колбочки. Предполагаемый принцип работы колбочки (её внешней конуснй мебраны) в режиме конусного волновода помещённого в среду прозрачного тела глаза сводится также к принципу оппонентного отбор основных квантованных спектральных лучей света не в цвете (рецепторный уровень). Например, самый совершенный цифровой фотоаппарат, колориметр, которые предназначены для получения цвета при воздействии на их фотодатчики, примерно как у глаза, на рецепторном уровне фиксируют оптическое изображение каждой предметной точки не цветное. Главное отличие состоит в том, что не цветное оптическое изображение в системе RAW (с без цвета) и в оптических приборах воспринимается плоским фотоприёмником — фотосенсором или фототранзистором. Например, фотосенсор, состоящий из мозаики ячеек трёх, четырёх пикселей, накрытых светофильтрами системы RGB содержит каждую клетку (блок) из "красного", "зелёного", "синего" пиксела (система RGB — трихроматизма), которая фиксирует сфокусированную предметную точку в виде трёх основных спектральных отфильтрованных лучей (трихроматизма) с квантованной энергией, равной длине волны, в виде заряда конденсатора-пикселя с определённым количеством электронов. Ещё с той разницей, что каждая сфокусированная предметная точка оценивается с полным набором её спектральных лучей тремя клетками, несущие информацию трёх рядом расположенных сфокусированных предметных точек по одному поглощённому спектральному лучу (набора RGB). Каждая предметная точка, сфокусированная на клетку, где каждый пиксель отбирает один основной спектральный луч из трёх падающих на него — или красный, или зелёный, или синий, фиксирует энергию длины волны одного из трёх лучей. Т.е. 1/3 цветовой информации каждой сфокусированной предметной точки. Для создания цветного изображения, в результате полученных данных, применяется АЦП, который формирует из полученных сигналов цветное оптическое изображение, но не аналоговое. (См. Аналоговая фотография).
Как видим, биологическая система зрения при формировании оптического изображения на чёрно-белом и цветном уровнях у нас происходят в отличие от научнотехнических достижений (цветная фотография) по законам биофизическим и биохимическим и имеют общие сходства, но и принципиальные отличия от электронно-оптических систем.
- Главные отличия:
- 1) Оптическое изображение в нашем сознании — аналоговое, индивидуальное и для нас является эталоном, согласно которому мы сравниваем со всеми изображениями, которые нас окружают, или которые мы создаём сами.
- 2) Наш фотоприёмник — сетчатка глаза с жёлтым пятном с центральной ямкой, где формируется 50% объектов цветного зрения.
- 3) Главным принципиальным элементом работы билогического фотоприёмка согласно данным учёных в настоящее время является колбочка, которая рассматривается как клетка с внешней долей конусной мембраны, имеющей пространственную структуру; и работающей по типу конусного волновода в среде прозрачного тела глаза. Это даёт возможность ей воспринимать сфокусированные лучи предметной точки оптического изображения в очерёдности согласно принципа работы конусного волновода, т.е. очерёдности восприятия падающих на него спектральтных лучей в виде порядка красные→зелёные→синие→оранжевые и т.д., при этом оппонентно отбирать основные спектральные лучи одной сфокусированной предметной точки (S,M,L,) или RGB не в цвете (внешняя мембрана — биологический конусный волновод внешней доли конусной мембраны колбочки).
Если для получения цветного изображения фотокамерой применяется фотосенсор, где клетка из пикселей RGB фильтрует только одну составляющую цвета сфокусированной предметной точки, т.е.1/3, то для получения всех трёх цветов при помощи АЦП из трёх клеток фотосенсора (трёх предметных точек в каждой, рядом расположенных) формируется цветное изображение не аналоговое, но ещё более красочное, чем оригинал. В мозгу же, на основании отобранных сигналов более коротким путём, при этом аналоговых сигналов каждой предметной точки, формируется полноценное наше аналоговое оптическое изображение.
- 4) Каждая колбочка в своих аксонах содержит пигменты кон-опсины: красные, зелёные, синие, которые участвуют в формировании, фототрансдукции оппонентном отборе биосигналов захваченных лучей предметной точки изображения. Отобранные основные спектральныи лучи с характеристиками каждой сфокусированной предметной точки) происходят на нейронном уровне, не в цвете. После такого отбора сигнала в мембране колбочки при сравнении трёх пар сигналов красный-зелёный, синий-жёлтый, чёрный-белый он отправляется по нервным стволам в зрительные отделы головного мозга. (См. Теория оппонентного цветного зрения).
Отличающиеся особенности колбочек[править | править код]
Отличающиеся особенности колбочек состоят в том, что они все одинаковые мофологически, но в зависимости от места расположения на сетчатке имеют отличающиеся размеры. Это связано с функцией их работы. Например, колбочки в центральной ямке имеют большую плотность расположения и воспринимают в основном красно-зелёные лучи имеют больший размер основания конуса внешней доли мембраны, т.к. поперечное сечения фронтального луча красного цвета наибольшее и соответсвенно основание конуса мембраны наибольшее, чем у синей колбочки. (Работа мембраны колбочек и палочек как волновод). С другой стороны все колбочки содержат разновидности фотопигмента кон-опсины, которые в зависимости от захваченного луча света предметной точки и оппонентного отбора выделения наиболее яркого основного луча триады RGB (S,M,L) в мембране выделяется фотопигмент кон-опсин данного цвета. У колбочек три разновидности фотопигмента йодопсина — фотопигменты хлоролаб и эритролаб. Фотопигмент хлоролаб поглощает лучи жёлто-зелёной части спектра света. Фотопигмент эритролаб поглощает лучи жёлто-красной части спектра. Третий вид фотопигмента «цианолаб» (предполагаемый) поглощает синие лучи света, который проявляется при захвате синих лучей колбочкой-S. Колбочки-S располагаются вне зоны ямки, в периферийной зоне сетчатки. Однако пока известно, что йодопсин находится во всех колбочках глаза.
Современный взгляд на работу колбочек[править | править код]
В 2009-2011 годах учёные физики, доктор физик Джеральд К. Хат[34] (см. Участник:Миг/Центральная ямка сетчатки глаза), учёный физик Джон А. Мейдерос[4] (см. Участник:Миг/Работа мембраны колбочек и палочек как волновод) по новому интерпретировали основы цветного зрения с точки зрения физики, которая не только не противоречит, а даже уточняет некоторые полученные выводы, данные известных учёных, предшественников — биологов, биофизиков, биохимиков.
Сейчас всё больше учёные рассматривают колбочку как клетку с внешней долей конусной мембраны, имеющей пространственную структуру, работающей по типу биологического конусного волновода в среде прозрачного тела глаза. Это даёт возможность колбочке воспринимать сфокусированные лучи предметной точки оптического изображения в очерёдности, как работает конусный волновод в прозрачном теле (жидкой среде) сетчатки глаза, в очерёдности (обратной): красные→зелёные→синие→оранжевые и т.д. как работают обычные волноводы в воздухе. При этом идёт отбор основныех спектральных квантованных лучей (S,M.,L,) или RGB, притом не в цвете (рецепторный уровень). Эти отфильтрованные квантованные лучи в виде длин волн предметной точки со своими характеристиками преобразуются, трансформируется со скоростью в фемтосекуду, а затем трансдукцируются в мозг, где сфокусированная квантованная предмтная точка оптического изображения (в квантах энергии её длины волны) не в цвете, сканируется со своими координатами, но уже в цвете. Сигналы в зрительных отделах головного мозга преобразовываются у нас в в виде аналогового оптического изображения в цвете, стерео. Это бинокулярное оптическое изображение (субъективное), которое создаётся у каждого из нас своё.
Никакой заменитель глаза не способен это оптическое изображение повторить, мы можем с применением современной техники приблизиться к глазу, познать его принципы работы, но в настоящее время существует много непознанных вопросов получения оптического изображения на сетчатке, в мозгу. Самое важное, созданная техника не может работать как пространственный биологический фоторецептор — конусная внешняя доля мембраны колбочки. Предполагаемый принцип работы колбочки (её внешней конуснй мебраны) в режиме конусного волновода помещённого в среду прозрачного тела глаза сводится также к принципу оппонентного отбор основных квантованных спектральных лучей света не в цвете (рецепторный уровень). Например, самый совершенный цифровой фотоаппарат, колориметр, которые предназначены для получения цвета при воздействии на их фотодатчики, примерно как у глаза, на рецепторном уровне фиксируют оптическое изображение каждой предметной точки не цветное. Главное отличие состоит в том, что не цветное оптическое изображение в системе RAW (без цвета). В оптических приборах изображение предметной точки воспринимается плоским фотоприёмником — фотосенсором или фототранзистором. Например, фотосенсор, состоящий из мозаики ячеек трёх, четырёх пикселей, накрытых светофильтрами системы RGB содержит каждую ячейку (блок) из "красного", "зелёного", "синего" пиксела (система RGB — трихроматизма), которая фиксирует напрямую сфокусированную предметную точку в виде трёх основных спектральных отфильтрованных лучей (трихроматизма) с квантованной энергией, равной длине волны, в виде заряда конденсатора-пикселя с определённым количеством электронов. Ещё с той разницей, что каждая сфокусированная предметная точка оценивается с полным набором её спектральных лучей тремя ячейками, несущие информацию трёх рядом расположенных сфокусированных предметных точек по одному поглощённому спектральному лучу (набора RGB). Каждая предметная точка, сфокусированная на клетку, где каждый пиксель фиксирует один основной спектральный луч из трёх падающих на него — или красный, или зелёный, или синий, фиксирует энергию длины волны одного из трёх лучей. Т.е. 1/3 цветовой информации каждой сфокусированной предметной точки. Для создания цветного изображения, в результате полученных данных, применяется АЦП, который формирует из полученных сигналов цветное оптическое изображение, но не аналоговое. (См. Аналоговая фотография).
Как видим, биологическая же система зрения формирования оптического изображения на чёрно-белом и цветном уровнях у нас происходят в отличие от физикотехнических достижений (цветная фотография) по законам биофизическим и биохимическим и имеют общие сходства, но и принципиальные отличия от электронно-оптических систем.
- Главные отличия:
- 1)Оптическое изображение в нашем сознании — аналоговое, индивидуальное и для нас является эталоном, согласно которому мы сравниваем со всеми изображениями, которые нас окружают, или которые мы создаём сами.
- 2) Наш фотоприёмник — сетчатка глаза с жёлтым пятном с центральной ямкой, где формируется 50% объектов цветного зрения.
- 3)Главным принципиальным элементом работы билогического фотоприёмка согласно данным учёных в настоящее время является колбочка, которая рассматривается как клетка с внешней долей конусной мембраны, имеющей пространственную структуру; и работающей по типу конусного волновода в среде прозрачного тела глаза. Это даёт возможность ей воспринимать сфокусированные лучи предметной точки оптического изображения в очерёдности согласно принципа работы конусного волновода, т.е. очерёдности восприятия падающих на него спектральтных лучей в виде порядка красные→зелёные→синие→оранжевые и т.д., и самое важное при этом оппонентно отбирать основные спектральные лучи одной сфокусированной предметной точки (S,M,L,) или RGB не в цвете (внешняя мембрана — биологический конусный волновод внешней доли конусной мембраны колбочки).
Если для получения цветного изображения фотокамерой применяется фотосенсор, где ячейка из пикселей RGB фильтрует только одну составляющую цвета сфокусированной предметной точки, т.е.1/3, то для получения всех трёх цветов при помощи АЦП из трёх клеток фотосенсора (трёх предметных точек в каждой, рядом расположенных) формируется цветное изображение не аналоговое, и даже ещё более красочное, чем оригинал. В мозгу же, на основании оппонентно отобранных сигналов более коротким путём, аналоговых сигналов каждой предметной точки, формируется полноценное наше аналоговое оптическое изображение.
- 4)Каждая колбочка в своих аксонах содержит пигменты кон-опсины: красные, зелёные, синие, которые участвуют в формировании, фототрансдукции оппонентном отборе биосигналов захваченных лучей предметной точки изображения. Отобранные основные спектральныи лучи с характеристиками каждой сфокусированной предметной точки) происходят на рецепторном уровне, не в цвете. После такого отбора сигнала в мембране колбочки при сравнении трёх пар сигналов красный-зелёный, синий-жёлтый, чёрный-белый он отправляется по нервным стволам в зрительные отделы головного мозга. (Нейронный уровень). (См. Теория оппонентного цветного зрения).
Особенности работы S-колбочек[править | править код]

- Положение колец (см. рис. R) — длинной длины волны (L), чувствительной к красному цвету в центральном месте ямки. Центр (ямки)(550нм) — середина чувствительности полосы в 7-8° (величина угла конусный поверхности в степенях), где колбочки находятся вне окружения палочками с углом зоны «cone peak» 1° (рис. 1г). Колбочки окружены палочками с короткими длинами волны и взаимодействующими за пределами конусного угла 20° (степеней). Постоянно об этом положении колбочек и палочек традиционно думали как отклонение, вызванное продольной (осевой) хроматической аберрацией глаза. И такая работа демонстрировала бы, что этот ответ — не отклонение вообще, а скорее фундаментальное основание для обработки изображения глаза. В этом контексте доктор Huth отметил комментарий Millidotа, показанного ранее. При этом, имея критически точную, опорную базовую длину волны (midband длину) 550нм, скорости формирования сигналов сфокусированной предметной точки в фемтосекунду, например, синего сигнала S, создают условия в фовеальной ямке для нахождения величины длины волны синего цвета (см. рис.1p) S-колбочками, которые к ним направляются из зоны центральной ямки при взаимодействии колбочек M/L. (Cм. рис. R).

Например, демонстрировалось, что горизонтальные клетки производят отрицательную обратную связь с колбочками (Baylor и др., 1971; Fuortes и Саймон, 1974; Burkhardt, 1993), которые заняты в центре, и окружают поколение в биполярных клетках. Они красят ответы противника в горизонтальных клетках животных с хорошим цветным видением (Fourtes и Саймон 1974). В случае данных объекта ПРИВЕТ и работы горизонтальных клеток HII сетчатки примата. Это было трудным исследованием к демонстрируемому цветному сопротивлению во внутриклеточной регистрации и маркировках краски (Dacey и др., 1996), хотя анатомически известно, что есть цветная селективность их контактов (Ahnelt и Kolb, 1994). Dacey и соавторы (1996) заключили, что клетки HII, которые конечно вступают в контакт с L-и М. колбочками также, но прежде всего которые заинтересованы в конусах (S-колбочках), которые не должны участвовать в сигналах обратной связи с более длинными конусными мембранами S-колбочек. Однако, возможно что обратная связь клеток HII относится только к Булочкам, формируя таким образом сопротивление конусах непосредственно с их биполярными клетками. Это должно привести к Булочке противника биполярного СЫНА, при закрытом L-/M-OFF (синий НА/жёлтый цвет ПРОЧЬ) канале, который можно было бы нести прямо к клеткам нервного узла (см. рис.9). Это значит, что сфокусированные синие монолучи предметной точки на сетчатку отбираются предварительно биполярными клетками из красно-зелёных, жёлтых монолучей, перед передачей их S-колбочке для фототрансдукции синего сигнала в мозг[36].

И как показано в диаграмме «Розетты Стоун» выше, (geoemetric) геометрическое строительство, используемое здесь определяет точный центр — 550 нм — визуальной полосы и, кроме того, точное место, где эта длина волны взаимодействует в пункте полной восьмугольной симметрии в относящейся к сетчатке глаза оригинальности конуснонго 7-8° (степеней). Приняв данный случай получаем, что это обеспечивает «неподвижную ссылку длины волны» (расположение в центре базисной длины волны 550нм), которую предлагает Доктор Huth, и служит основанием для цветного постоянства видения и синтеза оттенков, что мы называем цвет, как описано Эдвином Ландом. Все мы приходим к тому же самому цвету, потому что, в основном, все мы имеем тот же самый размер/диаметр фоторецепторов. Здесь снимаются вопросы хроматической аберрации, фильтрации основных лучей, наличия необходимых пигментов, определения в сетчатке только трёх, геометрически-определенных длин волны. Мы приходим к обнаружению среди них максимальных границ, исключительно, длин волн: от длинной волны до короткой (700нм и 400нм) визуальной полосы, что даёт критически точное, геометрически определённую опорную, базовую (midband) длину волны (550 нм) для нахождения величины длины волны синего цвета (S).






Дифференцированная работа на фокальной поверхности сетчатки фотодатчиков — экстерорецепторов[править | править код]
На основании последних данных (См. рис. 1,1а,1в,1г, доктор Джеральд К. Хат [44] и доктор John A. Medeiros [4]учёные вообще, хоть и немноо гипотетически, на базе всего набора информации по цветному зрению, более реалистично пришли к обобщению, что цветное зрение следует рассматривать с точки зрения дифференцированной работы на фокальной поверхности сетчатки фотодатчиков в виде нано-антен или структур колбочка в окружении шести или восьми палочек.Каждая нано-атена - пространственная структура, что согласуется с тезисом, что каждая колбочка работает как конусный волновод в среде прозрачного тела глаза. Данное направление снимает вопросы хроматической аберрации, фильтрации основных лучей, наличия необходимых пигментов, определения в сетчатке только трёх, геометрически-определенных длин волны и обнаружение среди них максимальных границ, исключительно длин волн: от длинной волны до короткой (700нм и 400нм) визуальной полосы и, даёт критически точное, геометрически определённую опорную, базовую (midband) длину волны (550 нм) для нахождения величины длины волны синего цвета (S).
Например, подтверждая вышесказанное, в работе Helgе Kolbа дано:
- Электронная микроскопия, наконец, показала, что тип HII горизонтальной клетки действительно послал много древовидных «процессов» (сигналов) немногим конусам (колбочки S) посредством её древовидных полевых и меньших концентрациях процессов, приведших к лежанию "M." и L-колбочек. Короткие аксоны этих клеток HII связываются с конусами исключительно как на рис. 8b,9 (Ahnelt и Kolb, 1994). Внутриклеточная регистрация от горизонтальных клеток H2 в сетчатке обезьяны доказала окончательно, что эта горизонтальная синяя клетка — чувствительный и важный элемент тропы конуса в сетчатке примата (Dacey и др., 1996)[36].
Процесс фототрансдукции[править | править код]
Процесс фототрансдукции — важный и сложный процесс, объясняющий механизм поглощения фотонов, преобразования и передачи электрических биосигналов фотодатчиками. И чтобы понять его, нужно иметь представление о структуре клеток фоторецепторов, вовлеченных в зрительный процесс: о палочках, колбочках и клеток внутренних слоёв сетчатки. Клетки колбочек и палочек содержат хромофор, (11-cis-retinal, Альдегид Витамина A1[45] и легко-абсорбирующей части) обязанный белку мембраны клетке опсину (opsin). Палочки имеют дело со слабым освещением (низким сумеречным уровнем) и не добиваются цветного видения. Колбочки, с другой стороны, могут закодировать цвет изображения через сравнение продукций трёх различных типов колбочек! Каждый тип колбочки лучше всего отвечает на определенные длины волны, или цвета, света, потому что каждый тип имеет немного различный опсин (opsin). Три типа фоторецепторов: L-колбочки, М-колбочки и Булочки оптимально отвечают на длинные длины волны (красноватый цвет), средние длины волны (зеленоватый цвет), и короткие длины волны (синеватый цвет) соответственно[45][46]. Амакриновые клетки играют важную роль в вопросах регулирования и передачи сигналов колбочек и палочек в мозг. Они образуют чрезвычайно сложную, соединенную щелевыми контактами сеть, а отросток, не покидая пределов сетчатки, ветвится в области синаптических контактов биполярных ганглиозных нейронов, они изменяют характер распространения возбуждения сигналов по нервным клеткам ганглиозного слоя.
Фоторецепторы и фотопигменты сетчатки[править | править код]
| Клетки сетчатки Нейронов, Палочек, Колбочек, ipRGC | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

(Клетки при большом увеличении). | ||||||||||||
| ||||||||||||
Фоточувствительные экстерорецепторы сетчатки — экстерорецепторы, расположенные в сетчтаке Глаза, воспринимающие электромагнитное излучение в световом диапазоне длин волн. Фоторецепторы обеспечивают и создание оптического изображения — функцию Зрения, и внешнюю коррекцию биоритмов сна и бодрствования, зависящую от общей освещённости.
Это группа специализированных светочувствительных образований в сетчатке, воспринимающих свет и по-разному возбуждающихся от воздейстаия них световых лучей, с ответом в виде фототрансдукции (передачи «преобразованного» сигнала). Фоторецепторы создают т. н. потенциалы действия — нервные импульсы, дополнительно обрабатываемые в зрительной коре головного мозга, при помощи совокупного движения, поведения биологических систем, которые замкнуты и одновременно имеющие модулятор движения (рефлексы), связывающий фоторецепторы и всех его внутренних элементов с движением внешней среды (электромагнитнх волн), из которой поступает движущийся сигнал, и регулирующее по амплитуде или частоте движение саморегуляции биологических систем. Например, регулировка функции фочувствительного элемента фоторецепторов (мембран колбочек, палочек) означает автоматическую поднастройку оптимального положения их при раздражении падающими фокусирующимся основными спектральными лучами RGB. При этом диспергированный «белый» луч света должен иметь как минимум семь спектральных лучей фокусировки спектра вдоль оси мембраны колбочек на разной высоте, расположенных в блоках. Каждый блок имеет диаметр примерно 7мкм (совпадающий с площадью, сфокусированного блока двух предметных точек с просветом на расстоянии 250мм размером 0,176мм - разрешаюшая способность нормальной зрительной системы человека с остротой зрения 1,0 (см. Участник:Миг/Острота зрения)). Каждый монохроматический луч фокусируется в зоне конуса мембраны с диаметром основания в 3-5мкм и высотой = 50мкм. В блоке может разместиться примерно 3-6 колбочек, что достаточно с точки зрения теории оппонентности Эвальда Геринга — обработки цветового воздействия. Т.е. спектральные монохроматические лучи любой предметной точки (например, 0,176мм) изображения фокусируются в виде кружков нерезкости диаметром ~7мкм и улавливаются как бы рефлекторно колеблющейся, самонастривающейся, подвижной системой колбочек с фоторецепторами (палочками и колбочками), чувствительными к своему цвету[47][48]
Эволюция и генетика фотопигментов[править | править код]
Развитие и генетика фотопигментов — направление исследований о развитии и генетике фотопигментов, связанной с наличием аутосомальных генов, которые производят фотопигменты: колбочки-S и X-связанные пигменты с пиковой чувствительностью в зелёном и красном диапазоне длин волн — пигмент M/L.
Экстерорецепторы[править | править код]

Слева — микрофотография сечения сетчатки глаза рыбы, справа — схематическая прорисовка взаиморасположения органелл и фоторецепторов. Направление света, приходящего в сетчатку сквозь хрусталик: снизу-вверх.
На фото видно: днём палочки скрыты гранулами меланина; рецепторная часть колбочек выдвинута к свету (на фото — смещена вниз).[49]

Справа — схема перемещений органелл и изменения формы фоторецепторных клеток.
На фото видно: при недостатке света гранулы меланина открыли путь света к палочкам; рецепторная часть колбочек удалена от источника света, рецепторная часть палочек — приближена к нему (на фото — смещена вниз).[50]
Ретиномоторная реакция фоторецепторов сетчатки глаза — механические процессы в сетчатке глаза, связанные с перестройкой взаимного расположения фоторецепторов (палочек и колбочек), и гранул меланина,
в соответствии с уровнем освещённости (см. рис. 1/1-2/2), где видно, что колбочки работают при дневном освещении без участия палочек, а палочки работают при сумеречном и ночном освещении без колбочек! (Это важное открытие).
Экстерорецепторы ipRGC[править | править код]
Третий вид фоточувствительных клеток (см. Сечение сетчатки глаза) (не колбочки и не палочки), которые относятся к сетчатке глазаа[Wong, Kwoon Y.; Dunn, Felice A.; Berson, David M. (22 декабря, 2005). "«Photoreceptor Adaptation in Intrinsically Photosensitive Retinal Ganglion Cells» (HTML: Full text)" 48. Neuron. pp. 1001—1010. doi:10.1016/j.neuron.2005.11.016. Retrieved 11 мая, 2011. Check date values in: |date=, |accessdate= (help)].
- pRGC (Retinal ganglion cell) — все фоточувствительные нервные клетки;
- ipRGC — фоточувствительные нервные клетки D,E, относящиеся к сетчатке глаза. (Фоточувствительные клетки нервного узла ipRGC содержат фотопигмент меланопсин[2] и расположены в сетчатке вне фокальной поверхности в слое Ganglionic layer (см. Сечение сетчатки глаза).)
Клетки нервного узла (ipRGC), содержащие пигмент (melanopsin)[3], — клетки, которые недавно обнаружены и являются типом нервной клетки в сетчатке [4] глаз млекопитающих. В отличие от других нервных узлов [5], относящихся к сетчатке глаза, они являются фотосенсибилизационными Фотосенсибилизация!. Это означает, что они — третий класс фоторецепторов сетчатки глаза, которые возбуждаются под действием лучей света. (Они даже при блокировании классических фоторецепторов палочек и колбочек, или в случае применения фармакологических агентов их блокировки, или при отделении их от сетчатки, независимо функционируют), т.е участвуют в зрительном процессе.
Палочки[править | править код]
Па́лочки англ. rod cells) — один из типов фоторецепторов, расположенных в фокальной поверхности сетчатки в слое (экстерорецепторов), периферических отростков светочувствительных клеток сетчатки глаза, названный так за свою цилиндрическую форму. Это высокоспециализированные клетки, преобразующие световые раздражения в нервное возбуждение (сигналы).
Палочки чувствительны к свету благодаря наличию в них специфического пигмента — родопсина (или зрительный пурпур).
Колбочки[править | править код]
Ко́лбочки (см.рис.1) — (англ. cone) один из типов фоторецепторов (экстерорецепторы), расположенных в фокальной поверхности сетчатки в слое (экстерорецепторов), периферических отростков светочувствительных клеток сетчатки глаза, названный так за свою коническую форму. Это высокоспециализированные клетки, воспринимающие и преобразующие световые сигналы (фотоны) в нервное возбуждение в виде биоэлектрических (сигналов) на рецепторном уровне (есть уровень нейронный — участие в работе зрительных отделов головного мозга) на базе принципа оппонентного отбора более ярких сигналов.
Колбочки чувствительны к свету благодаря наличию в них специфического пигмента — йодопсина.
Головной мозг и оптическое изображение[править | править код]

Зрительные отделы головного мозга — восприятие цвета и света, получение оптического изображения в коре головного мозга — второй, окончательный этап работы зрительной системы образования оптического изображения в зрительных отделах головного мозга (см. рис.9).
Цветное зрение у человека и приматов[править | править код]

Механизмы трехцветного световосприятия у приматов (Trichromatic) лучше всего исследованы на примере человека. Согласно последним достижениям в исследованиях физиологии зрительной системы животного мира, у остальных приматов Участник:Миг/Зрение незначительно отличается от восприятия цвета человеком.
Следует заметить, что у части людей, вследствие определённой мутации, возник дополнительный зрительный пигмент, с несколько иным строением и иной спектральной чувствительностью. У таких людей (а пока известно, что эта мутация передаётся только женщинам) зрение четырёхкомпонентно, т.е. они несколько лучше различают оттенки различных цветов.
Цветное зрение у людей и животных связано с вопросами саморегуляции организма, с наличием совокупного движения, поведения биологических систем, которые должны быть замкнутыми и одновременно должны иметь модулятор движения, связывающий организм и его внутренние системы с подвижностью, колебаниями внешней среды, из которой поступают сигналы, воздействующие на элементы нервных узлов. Например, воздействуя на пигмент миоид сократимых фибрил колбочковых клеток происходит изменение высоты колбочки, что влечёт к изменению положения элементов нервной клетки фоторецепторов мембран, что означает возможную поднастройку положения их по высоте при раздражении падающимми, фокусирующимся спектральными лучами RGB. Здесь диспергированный «белый» луч света должен иметь как минимум семь спектральных точек фокусировки семи лучей спектра на оси мембраны на разной высоте с площадями кружка нерезкости около 7-9мкм, при этом на семи уровнях (зона цилиндра кружков нерезкости, где может разместиться ячейка как минимум из 6-ти колбочек). Это достаточно с точки зрения теории оппонентности Эвальда Геринга — отбор и обработки сигналов цветового воздействия на экстерорецепторы. Т.е. спектральные лучи любой точки оптического изображения фокусируются в виде кружков нерезкости диаметром 7-9мкм и улавливаются как бы рефлекторно колеблющейся, самонастривающейся, подвижной системой ячеек мембран экстерорецепторов (палочек и колбочек), чувствительных к своему цвету. Данные механизмы световосприятия с точки зрения теории оппонентности Эвальда Геринга — отбора и обработки сигналов цветового воздействия на экстерорецепторы (1985 г) согласуются данными рентгеноскопии (1977 г.) и флюоресцентной наноскопии (2007 г.), которые однозначно фиксируют работу мозаики блоков колбочек RGB сетчатки, а также независимую работу палочек в условиях слабого освещения. При этом всё это происходит при непосредственном взаимодействии с фоторецептором ганглиозного слоя ipRGC сетчатки глаза в механизме восприятия зоны видимых сине-фиолетовых монолучей (см. ниже Фиг. R,К).
Трихроматизм и принцип оппонентности[править | править код]


Визуальное цветное зрение — основа современных теорий цветного зрения. Цветовое зрение, происходящее в зрительной системе инициируется поглощением света с помощью трех различных спектральных классов "шишек" (колбочек). Следовательно, цветовое видение описывается как трёхвариантное восприятие осноных цветов или как восприятие, ощущение цвета. Первоначально психофизические исследования показали, что цвета могут быть настроены на использование трех различных системах (праймериз). В 1802 году, Томас Молодых предложил модель, по которой восприятие цвета может быть закодировано на три основных цвета фоторецептоов, но не на кодировании тысяч цветовых рецепторов для отдельных цветов.[51]
Принцип оппонентного цветного зрения[править | править код]
В то время как теория трихроматизма [6] определяет способ, которым сетчатка глаза позволяет визуальной системе обнаруживать цвет с тремя типами колбочек, теория процесса противника (Теория оппонентного цветного зрения) является механизмами, которые получают и обрабатывают информацию от колбочек. Хотя трихроматизм и оппонентные процессы противника, как первоначально думали, имели разногласия, это было вначале, но чтобы это стало понятым требуется принять, что механизмы, ответственные за процесс противника, получают сигналы от трех типов колбочек и обрабатывают их на более сложном уровне[52].
Помимо колбочек, которые обнаруживают свет, входящий в глаз, биологическая основа теории противника вовлекает два других типа клеток: биполярные клетки, и клетки нервного узла. Информацию от колбочек передают к биполярным клеткам[53] (Биполярная клетка - тип нейрона, который имеет два расширения и которая как клетка сетчатки служит, чтобы управлять мускулами и передавать выходной (двигатель) зрительный сигнал в зрительные отделы головного мозга из сетчатки. Они являются ячейками в оппонентном процессе противника, которые преобразовывают информацию от колбочек и передают информацию к клеткам нервного узла, которые представляют два главных класса клеток: magnocellular [7], или слои большой клетки, и parvocellular [8], а также представляют слои маленькой клетки. Клетки Parvocellular, или клетки P, работают в основном с большей частью информации о цвете, и его попадании на сетчатку в виде двух групп: той, которая обрабатывает информацию о различиях между L и М колбочками при отборе, и той, которая обрабатывает различия между колбочками S и объединенным сигналом, а также и от L и от М. колбочек. Первый подтип клеток ответственен за обработку красно-зеленых различий сигналов, и второй тип — за синие-желтые различия. P клетки также передают информацию об интенсивности света (выбор более ярких цветов), в зависимомости от их восприимчивости в этих областях (см. Зрительные отделы головного мозга). [54]
Цветное зрение у птиц[править | править код]
Доказано[5] (2006 год), что сетчатка у птиц обладает одной из самых сложных систем фоторецептора колбочки среди позвоночных животных. Исследуя спектры поглощения сетчатки было получено, что птицы имеют пять типов колбочек, включая четыре одиночных, которые поддерживают видение цвета четырехроматизма(tetrachromatic) и двойную (колбочку), которая обеспечивает восприятие движения. (См. рис. 2a).
Когда пигмент поглощает свет, падающий на сетчатку глаза (на палочки и колбочки), изменяется мембранный потенциал клетки-колбочки, затрагивающий нейроны в слое ганглий сетчатки. Каждый нейрон в слое нервного узла может воспринимать информацию процессов от множества фоторецепторов, и может в свою очередь вызвать нервный импульс и передать информацию по оптическому нерву для дальнейшей обработки в специализированные зрительные центры мозга. Чем более интенсивный свет, тем больше фотонов поглощено зрительными пигментами, и большее возбуждение каждой клетки стимулирует появление более яркого оптического изображения[5]. При этом предварительно монохроматические диспергированные лучи перед попаданием в мембрану попадают в жировые капелки, где оппонентно фильтруются наиболее сильные, яркие сигналы, которые в результаце фототрансдукции фильтрутся (согласно их цветовой специализации) и направляются по зрительным нервным стволам в мозг. С учётом оппонентного принципа отбора более яркого сигнала на этапе рецепторной работы зрительной системы выделение сигнала производится с участием ганглиозных слоев нейронов сетчатки, параллельно с выделением занимаются подавлением второстепенных, слабых сигналов, сжатием потоков из 6млн. до 1,2млн. для передачи по 1,2млн. нервных каналов. При этом с участием зрительных отделов головного мозга, связанного с ганглиозными фоторецепторами, происходит фильтрация трансдукцируемых цветных сигналов через фотосенсор ipRGC в прохождении фиолетово-синих лучей благодаря наличию у них фотопигмента меланопсина.
К тому же цвет жировых капелек каждой колбочки, ещё до попадания сигнала в мемрану, (например, синий, зелёный, красный) яркое доказательство трихроматизма цветного зрения. Т.е. на этапе уже рецепторном восприятие цвета происходит колбочками с разновидностью фотопигмента родопсина, которые отбирают свои яркие спектральные области монолучей (S,M.,L) предметных точек оптического изображения.
Математика цветного зрения[править | править код]
Выводы[править | править код]
Анализ данных цветовосприятия в последние 35-40 лет показывает, что c появлением новой техники, технологии исследований, учёным удалось доказать, что принцип трихроматизма (трёхкомпонетый принцип цветного зрения) является основой цветного зрения. На базе исследований на живых клетках при воздействии на них видимых лучей света (цвета) в цвете удалось рассмотреть колбочки S,M,L (RGB), которые участвуют в процессе дневного зрения, в процессе формирования цветного изображения в зрительных отделах головного мозга в трёхмерном пространстве (стерео) в цвете. C открытием третьего фоторецептора ганглиозного слоя сетчатки (см. ipRGC (~2007 год)), который расположен не в фокальной поверхности сетчатки, синаптически связан с колбочками, палочками, мозгом. Он выполняет важные функции при цветовосприятии в регулировании работы колбочек при восприятии спектра коротковолновых лучей (синих-фиолетовых), пропуская нормальные лучи спектра с длиной волны более 498нм, перенаправляя их в зону восприятия синих лучей колбочек-S , блокируя сильнодействующие синие лучи с высокой частой колебания — длинами волн менее 498нм.. Он содержит оптический пигмент меланопсин, чувствительный к синим лучам (S) света.
Проводились исследования учёными Е.О.Загальской и В.П.Гнюбкиной из Института Биологии Моря ДВО РАН (2006 год), Владивосток. Цель этого исследования состояла в том, чтобы проанализировать ретиномоторные движения у взрослых рыб (zebrafish) и процесс созревания ретиномоторной реакции в его развитии у молодняка рыб. Показано, что ретиномоторные движения используются как адаптация зрения в условиях тёмно/светового механизм адаптации у рыбы-зебры. У взрослых особей гранулы меланоцитов RPE

В 2006 [55] году при помощи трансмиссионной электронной микроскопии пигментного слоя сетчатки исследован эпителий клеток сетчатки глаза, где вырабатывается пигмент меланин (коричневого и чёрного цвета), который обеспечивает открытие и закрытие зон освещения колбочек и палочек в зависимости от вида освещения. При дневном освещении данный пигмент блокирует работу палочек, которые ко всему ещё уходят в зону под колбочки. И наоборот, при сумеречном освещении палочки выходят из под прикрытия зоны прохождения синих и ультрафиолетовых лучей меланином, а колбочки опускаются в зону ниже палочек и находятся под прикрытием меланина. (См. рис.C). В конечном итоге подтверждено открытие явления ретиномоторной реакции фоторецепторов, работы колбочек и палочек в условиях дневного и ночного освещения учёными Е.О.Загальской и В.П.Гнюбкиной из Института Биологии Моря ДВО РАН, Владивосток.
- Рентгеноскопия сетчатки и отделов мозга, проведенная ещё в 1966, повторно в 2009 годах Р.Е.Марком, на атомно-молекулярном уровне подтверждает ранее сформулированые гипотезы о работе сетчатки, где заложены фоторецепторы колбочки, выделющие основные лучи спектра S,M,L ("синие","зелёные", "красные") (см. Фиг.R,рис.S), которые на рецепторном уровне воспринимают, трансдукцируют основные сигналы RGB оптического изображения (контурного) перед передачей их в высшие зрительные отделы (нейронные) головного мозга (см.Лаборатория Р.Е.Марка). При этом следует подчеркнуть, что морфологически колбочки устроены одинаково, но в зависимости от места расположения они имеют разные размеры, например, в центральной ямке сетчатки они самые маленькие, т.к. плотно упакованы и при этом, в зоне ямочки диаметром до 1мм расположены только колбочки красные и зелёные, где нет палочек вообще. Каждая колбочка способна выделить один из сигналов S,M,L, т.к. фотопигмент опсин в зависимости от сфокусированного и выделенного основного луча предметной точи способен в результате мутации белка пигмента принимать красную, зелёную и синюю окраску!
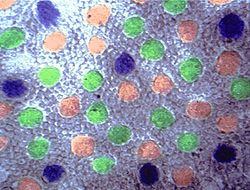 Рис.S. Распределение рецепторов в сетчатке бабуина . Синие колбочки были распределены регулярно в периферии, красные и зеленые колбочки были распределены беспорядочно всюду. Плотность распределения зелёных колбочек больше, чем красных, больше чем синих[16].
Рис.S. Распределение рецепторов в сетчатке бабуина . Синие колбочки были распределены регулярно в периферии, красные и зеленые колбочки были распределены беспорядочно всюду. Плотность распределения зелёных колбочек больше, чем красных, больше чем синих[16].

В настоящее время уже вообще не стоит вопрос о восприятии цвета в сетчатке блоками из трёх колбочек S,M,L (R.Marc, 1977)[16][56]. Если говорить о восприятии синих монолучей синими колбочками, то уже в 1977 году получены легко рентгеновские снимки синей колбочки в ряду основных колбочек зелёных и красных (см. рис. S).
Группа учёных Лаборатории R.Marcа (Лаборатория Р.Е.Марка) уже работают над созданием атласа основных нервных клеток сетчатки и отделов головного мозга, на основании электронной микроскопии их у кроликов и крыс. При этом используется разработаная мощная программа обработки данных современных микроскопов и компьютерных систем. При этом открывается возможность визуально анализировать работу здоровых и больных клеток с целью лечения целого ряда болезней (слепота, дальтонизм и др[16].
Что важно, на рентгеноскопии среза сетчатки (cм. Фиг.R) видно, что основные фоторецепторы колбочки и палочки в процессе формирования оптического изображения работают независимо. Так синяя колбочка при дневном зрении воспринимает диапазон синих лучей под присмотром фоторецепторов ipRGC. Например, сильные сигналы ультрафиолетовых, синих лучей под действием ipRGC и мозга закрывают вообще ресницами глаза глаз от их попадания во внутрь (скорость реакции 1/2000сек) и имеются другие функции у них, связанные с трансдукцией синего сигнала.
- Доводы о том, что палочки и колбочки одинаковы и не имеют различий голословны и оторваны от жизни. В настоящее время получены изображения разных типов колбочек и палочек, которые отличаются топографически и функционально. (См. рис. 11). На приведенных изображениях колбочек птиц и черепах ещё раз показан вид колбочек, которые морфологически одинаковы, но различаются видом, в зависимости от места расположения и меняющемуся виду при работе. В глазу птиц даже есть двуглавые колбочки, расположенные в периферийной части и управляющие движением птиц.
 Рис.11;Блок колбочек черепахи с цветными жировыми капельками[57]
Рис.11;Блок колбочек черепахи с цветными жировыми капельками[57]

 Рис.П; Мутация белка протеина (содержащего разновидность пигмента родопсин) в мембране колбочки человека в четырёх областяхЧто касается фотопигментов, то следует знать, что пигмент, например, фоторецептора или изменяют своё устройство или претерпевают уменьшение своего изображения (вида), когда они поглощают фотон. Это изменение происходит в устройстве или в окислительно-восстановительном процессе хромофора и выражается в изменении устройства белка или его деятельности, и вызывает каскад трансдукции сигнала. Примеры для пигментов фоторецепторов, находящиеся в сетчатке глаза: например, родопсин (относящийся к сложным белкам хромопротеинам) вызывает жёлтый цвет; например, криптохром (cryptochrome), и билин (bilin) вызывает голубой цвет растений. Относительно фотопигмента цианолаба, который ещё не выделен учёными, то в результате фототрансдукции сигнала, пигмент который светится в жировых капельках синим цветом увиден и его можно назвать любым именем, важен цвет, который виден при исследованиях на снимках, и который имеется на синих колбочках-S. Не исключено, что он близок к меланопсину, билину. В любом случае, он является разновидностью фотопигмента опсина (См. Белок и передача зрительных биосигналов) [цитата, необходимая]
Рис.П; Мутация белка протеина (содержащего разновидность пигмента родопсин) в мембране колбочки человека в четырёх областяхЧто касается фотопигментов, то следует знать, что пигмент, например, фоторецептора или изменяют своё устройство или претерпевают уменьшение своего изображения (вида), когда они поглощают фотон. Это изменение происходит в устройстве или в окислительно-восстановительном процессе хромофора и выражается в изменении устройства белка или его деятельности, и вызывает каскад трансдукции сигнала. Примеры для пигментов фоторецепторов, находящиеся в сетчатке глаза: например, родопсин (относящийся к сложным белкам хромопротеинам) вызывает жёлтый цвет; например, криптохром (cryptochrome), и билин (bilin) вызывает голубой цвет растений. Относительно фотопигмента цианолаба, который ещё не выделен учёными, то в результате фототрансдукции сигнала, пигмент который светится в жировых капельках синим цветом увиден и его можно назвать любым именем, важен цвет, который виден при исследованиях на снимках, и который имеется на синих колбочках-S. Не исключено, что он близок к меланопсину, билину. В любом случае, он является разновидностью фотопигмента опсина (См. Белок и передача зрительных биосигналов) [цитата, необходимая] Фиг.К; Фигура «синей» колбочки сетчатки примата при электронной микроскопии и её связи с ячейками ганглиозного слоя сетчатки [16].
Фиг.К; Фигура «синей» колбочки сетчатки примата при электронной микроскопии и её связи с ячейками ганглиозного слоя сетчатки [16].


Например, проведенное исследование модели протеина (2009) «bestropin-1» — модель белка bestropin-1 в мембране рецепторов (приспособленный от Milenkovik), но только в четырёх областях, расположенных в мембране колбочек сетчатки человека, что говорит о процессах мутации и клетки белка, что показано в цвете и обозначено в цвете. (См.рис.П). [58]
- По вопросу хроматической аберрации, визуального восприятия предметных точек, остроты зрения, о строении фокальной поверхности сетчатки (мозаика, блоки фоторецепторов) и др.
При учёте хроматических аберраций глаза учитывается высота мембраны колбочки в 50мкм. Любой сфокусированный монохроматический луч спектра может сфокусироваться на разной высоте мембраны даже если все колбочки строго одинаково расположены в фокальной поверхности сетчатки. Однако и в этом случае мембраны колбочек на изображении среза сетчатки расположены не на одной высоте. Палочки расположены расположены выше колбочек. Подробнее смотри выше раздел «Мозаика экстерорецепторов сетчатки». При этом вопрос аберраций света в глазе имеет фундаментальный смысл цветного зрения. Т.к. при выделении колбочками основных световых сигналов S,M,L в блоке, например, трёх колбочек, а сфокусированный луч предметной точки фокусируется в виде кружка нерезкости равной или больше кружка нерезкости (3-5мкм), то в режиме оппонентного отбора наиболее ярких монолучей, каждая колбочка выделяет один из трёх основных лучей S,M,L, которые посылаются в мозг. При этом каждый сфокусированный луч ложится в конусной мембране в сечениях в зависимости от его длины волны, т.е.поперечного сечения фронта волны. Например, синий луч проходит до наименьшего размера сечения конусной мембраны колбочки, т.е. у вершины конуса. Или в пределах всей длины мембраны в 50мкм, лучи фокусируются точно на разных высотах, что исключает погрешности хроматической аберрации и обеспечивает глазу давать сигналы в мозг без искажений. Откуда оптическая система глаза не требует от преломляющей системы хрусталика и зрачка устройства по типу современных анастигматов как фотографии. Где все точки фокусируются в одной плоскости.
Дополнительно цветное зрение у людей и животных связано с вопросами саморегуляции организма, с наличием совокупного движения, поведения биологических систем, которые должны быть замкнутыми и одновременно должны иметь модулятор движения, связывающий организм и его внутренние системы с подвижностью, колебаниями внешней среды, из которой поступает сигналы, возбуждающие по амплитуде или частоте, требующие взаимодействия с элементами нервных узлов. Это относится также и к сетчатке с фоторецепторами. Живая ткань не может быть статичной, колбочки также подвержены ретиномоторной реакции, что сказывается на восприятие, зрительный процесс в режиме дневного и сумеречного освещения (См. Ретиномоторная реакция фоторецепторов).
На основании флюоресцентной наноскопии срезов живой сетчатки доказано, что восприятие света и цвета основано на работе фоторецепторов сетчатки, состоящей из мозаики блоков («клеток») с разным количеством колбочек, которые воспринимают основные монохроматические лучи спектра света предметной точки индивидуально в зависимости от строения сетчатки. Например, у птиц мозаика сетчатки состоит из блоков, содержащих систему колбочек «четырехроматик» (четыре колбочки), у человека —«трихроматизм» (три колбочки). (см. рис.3а,6). Т.е. в каждом блоке мозаики сетчатки могут содержаться по три, четыре, пять и т.д. колбочек, воспринимающих и выделяющих оппонентно основные лучи S,M,L (RGB).
- Также на основании опубликованных данных исследований профессора департамента офтальмологии функциональной нейроанатомии сетчатки R.E.Marcа (2009), то уже в 1966 году при электронной микроскопии сетчатки полученные данные «синих» колбочек приматов (см. фиг.К)[16].(См.Лаборатория Р.Е.Марка).
Например, из условия наличия 6 млн. колбочек в жёлтом пятне (у человека), на площади 6 мм², которые воспринимают цвет, можно на базе известных данных показать, что одна колбочка не в состоянии выдать нужную информацию цветов, сфокусированной на сетчатку предметной точки. Известно, что разрешающая способность нормального глаза при чтении с расстояния 250 мм находится в пределах 0,072-0,200 мм и в зависимости от освещённости и индивидуума примем среднестатистическую величину оценок разрешающей способности оптических приборов, среднестатистических глупп взрослых людей, проходящих тестирование (водителей транспортных средств, военнослужащих и т.д.) с показателем 0,0896мм (При остроте зрения 0,8). Основное количество фоторецепторов желтого пятна в центре сетчатки (~6млн) имеет площадь ~ 5,6-6 мм², (т.е. оптическое изображение содержит 1000000 (1мгп) разных цветовых точек), где: расстояние между одноименными точками (фоторецепторами или «пикселами») равно примерно 3-4мкм (диаметральный наибольший размер основания конуса мембраны колбочки). Рассматриваемые визуально, например, две предметные точки в виде облачков, кружков нерезкости с просветом, равный диаметру кружка нерезкости, которые глаз чётко видит. (Более подробно см. Современное понимание механизмов цветного зрения).


Из условия остроты зрения глаза — способности рассмотреть две точки с просветом между ними на определённом расстоянии (например, при остроте зрения человека 1,0 оно равно 72,5мкм), одна колбочка этого сделать не может. Откуда всегда рассматривается две точки с просветом минимум двумя колбочками или палочками, которые образуют блоки, например, RGB мозаики сетчатки (См. Цветное зрение у птиц). Как показано выше, диспергированные монолучи предметной точки с диаметром кружка нерезкости примерно 7мкм накрывают три колбочки мин. одного блока с диаметром 6,9мкм (где диаметр мембраны колбочек примерно = 3-4мкм). Таким образом, в любом случае предметные точки участвуют в восприятии изображения блоком из трёх колбочек мозаики сетчатки. А это значит, что 6млн. колбочек жёлтого пятна объединены в 1,5-2млн. блоков RGB, которые оппонентно трансдукцируют выходные биосигналы красного, зелёного, синего цвета под управлением головного мозга и фоторецепторов ipRGC, попадающие в зрительные отделы головного мозга через жировые капельки и далее по нервным путям. (Около 1млн. стволов). При этом часть сигналов из 1,5-2млн. фильтруется, подавляется в блоках при трансдукции RGB под управлением головного мозга и фоторецепторов ipRGC. Принято, что все колбочки одинаковые, каждая колбочка работает в оппонентном отборе основных лучей спектра S,M,L (RGB) предметной точки, которые на неё сфокусированы. При этом наиболее ярких.
Ссылкой на источник в виде:
- Единственные результаты, так ни кем и не оспоренные на сегодняшний день, были получены группой учёных W. B. Marks, W. H. Dobelle, E. F. Mak Nichol. Их исследование подробно описано в работе «Visual Pigments of Single Primate Cones» W. B. Marks, W. H. Dobelle, E. F. Mak Nichol Department of Biophysics, Johns Hopkins University, Baltimore, Maryland 21218. Science, Vol. 143, p. 1182, 1964., и что при освещении только её узкополосным излучением (вырезанным из широкого непрерывного спектра), по спектру отражения можно определить именно ту область, которую поглощает колбочка (область чувствительности колбочки). Для этого использовался широкополосный спектр оптического излучения из которого монохроматором вырезался очень узкий участок спектра. Далее этот монохромный спектр фокусировался в точку размером меньше, чем геометрический размер самой колбочки. Этой точкой сканировали сетчатку живого глаза.
Как видим, источник, где проведенное исследование в 1964 году при сканировании сетчатки не учитывалась работа мозга и фоторецептора ipRGC (открыты в 2007 году) с фотопигментом меланопсином, которые дозируют, фильтруют область сильных фотонов коротковолновых монохроматических лучей (фиолетовых-синих). Поэтому не случайно не удалось в то время обнаружить колбочек, трансдукцирующих синие лучи света.
- По вопросу дефектов цветовосприятия.
Именно, с открытием третьего фоторецептора сетчатки ipRGC (2007) учёные получили возможность глубже вникнуть в природу отклонений в восприятии цвета зрительной системой. Не рассматривая в комплексе работу колбочек, мозга, фоторецепторов ipRGC, конечно, не возможно оценить цветное зрение. Только с позиций трихроматизма, трёхкомпонетной теории цветного зрения практически решаются все вопросы цветового восприятия.
См. также[править | править код]
- Цветное зрение у человека (авторская основного пространства участника Миг)
- Оптический тракт
- Наружное коленчатое тело
- Зрение человека
- Измерение цвета
- Фотопигмент
- Опсины (авторская основного пространства участника Миг)
- Зрительная фототрансдукция
- Зрительные отделы головного мозга
- Зрительная кора
- Острота зрения
- Участник:Миг/Современный взгляд на световосприятие и цветное зрение
- Теория оппонентного цветного зрения (авторская основного пространства участника Миг)
- Специфика работы мембраны колбочек
- Ретиномоторная реакция фоторецепторов (авторская основного пространства участника Миг)
- Ночное зрение
- Дневное зрение
- Зелёный луч
- Эффект Трокслера
Примечания[править | править код]
- ↑ http://prometheus.med.utah.edu/~marclab/Marc_Duanes_FNAR_20080815_layout.pdf
- ↑ http://www.ghuth.com/
- ↑ http://www.ghuth.com/
- ↑ а б в г д John A. Medeiros. "Color Vision: A New Understanding". Cones and Color.net. Retrieved 8 сентября, 2012. Check date values in:
|accessdate=(help) - ↑ а б в г Goldsmith, Timothy H (Июль 2006). (PDF). Scientific American. pp. 69–75 http://www.csulb.edu/labs/bcl/elab/avian%20vision_intro.pdf. Retrieved 8 сентября, 2012. Text "title«What birds see» (PDF)" ignored (help); Check date values in:
|date=, |accessdate=(help); Missing or empty|title=(help) - ↑ http://www.ghuth.com/
- ↑ http://www.ghuth.com/
- ↑ М. А. Островский. "ФОТОБИОЛОГИЧЕСКИЙ ПАРАДОКС ЗРЕНИЯ" (PDF). Retrieved 26 июля, 2012. Check date values in:
|accessdate=(help) - ↑ Тонквист Г. Аспекты цвета. Что они значат и как могут быть использованы // Проблема цвета в психологии / Отв. ред. А.А.Митькин, Н.Н.Корж. М.: Наука, 1993. С. 5-53. (С. 7)
- ↑ ГР Jacobs, Neitz М., Deegan JF, и Neitz J. Trichromatic окрашивают видение в Новых Мировых обезьянах. Nature. 382: 156†“158, 1996}}
- ↑ Neitz М. и Neitz J. Молекулярная генетика цветного видения и цветных дефектов видения. Арч Офталмол 118: 691†“700, 2000. [FreeВ FullВ Текст]
- ↑ http://physiologyonline.physiology.org/content/17/3/93.full
- ↑ S-Potentials and Horizontal Cells. — Webvision. — С. 688. — ISBN 5-02-000747-1о книге<ul><li>Регулярное выражение «ISBN» классифицировало значение «5020007471↑http://www.biology.yale.edu/facultystaff/goldsmithTH.html↑http://uuhsc.utah.edu/MoranEyeCenter/faculty/robert_e_marc_phd.htm↑абвгде"Seeingincolor".Prometheus.Retrieved8сентября» как недопустимое.</li> <!--br--><li>Регулярное выражение «ISBN» классифицировало значение «2012. Checkdatevaluesin:|accessdate=(help)↑HumanPhysiologyandMechanismsofDisease."`UNIQitem77QINU`"↑цвета↑ "Цветовое зрение". 14 августа» как недопустимое.</li> <!--br--><li>Регулярное выражение «ISBN» классифицировало значение «2009. Retrieved 26 Июля» как недопустимое.</li> <!--br--><li>Регулярное выражение «ISBN» классифицировало значение «2010. Check date values in: |date=» как недопустимое.</li> <!--br--><li>Регулярное выражение «ISBN» классифицировало значение «|accessdate= (help)↑ Michael H. Rowe. "Trichromatic Color Vision in Primates". Physiology Online. Retrieved 8 сентября» как недопустимое.</li> <!--br--><li>Регулярное выражение «ISBN» классифицировало значение «2012. Check date values in: |accessdate= (help)↑ http://webvision.med.utah.edu/HC.html» как недопустимое.</li></ul>
- ↑ http://webvision.med.utah.edu/
- ↑ «Signatures of selection and gene conversion associated with human color vision variation». — С. 363-75.о книге
- ↑ Caulfield HJ (17 апреля, 2006). "«Biological color vision inspires artificial color processing»". SPIE Newsroom. doi:10.1117/2.1200603.0099. Retrieved 8 сентября, 2012. Check date values in:
|date=, |accessdate=(help) - ↑ «Some women may see 100 million colors, thanks to their genes». — Post-Gazette.com.о книге
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/colo perception/
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ http://webvision.med.utah.edu/photo2.html
- ↑ Конус
- ↑ http://www.ghuth.com/
- ↑ Color Vision: A New Understanding.
- ↑ http://www.ghuth.com/
- ↑ http://www.ghuth.com/
- ↑ а б в Helga Kolb. "S-Cone Pathways". Retrieved 8 сентября, 2012. Check date values in:
|accessdate=(help) - ↑ http://www.ghuth.com/
- ↑ http://www.ghuth.com/
- ↑ http://www.ghuth.com/
- ↑ http://www.ghuth.com/
- ↑ http://www.ghuth.com/
- ↑ Gerald Huth. "Found! The First Elusive Blue Cone". Ghuth.com. Retrieved 8 сентября, 2012. Check date values in:
|accessdate=(help) - ↑ http://webvision.med.utah.edu/photo1.html
- ↑ http://www.ghuth.com/
- ↑ а б Vitamin A
- ↑ GPCR
- ↑ Рецептор
- ↑ Фоторецептор
- ↑ http://www.imls.uzh.ch/static/CMS_publications/neuhauss/literatur/pdf06/Hodel_AnatRec_06.pdf
- ↑ http://www.imls.uzh.ch/static/CMS_publications/neuhauss/literatur/pdf06/Hodel_AnatRec_06.pdf
- ↑ http://webvision.med.utah.edu/book/part-viii-gabac-receptors/color-perception/
- ↑ Principles of Neural Science, 4th ed.. — McGraw-Hill. — С. 577–80.о книге
- ↑ Биполярные нейроны
- ↑ http://en.wikipedia.org/wiki/Opponent_process
- ↑ http://eps.dvo.ru/bm/2006/1/pdf/bm-055-059.pdf
- ↑ Marc RE, Sperling HG. (22 апреля, 1977). "Chromatic organization of primate cones". Retrieved 8 сентября, 2012. Check date values in:
|date=, |accessdate=(help) - ↑ http://webvision.med.utah.edu/photo2.html
- ↑ http://dare.ubn.kun.nl/bitstream/2066/74414/1/74414.pdf





{kind=link}
| Цвета и оттенки | |||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 |
|  | |||||||||||||||||||||||||||||||||||
| ∘ ∘ ∘ | |||||||||||||||||||||||||||||||||||||